|
x |
x |
|
 |
 |
|
INFECTIOUS
DISEASE |
BACTERIOLOGY |
IMMUNOLOGY |
MYCOLOGY |
PARASITOLOGY |
VIROLOGY |
|
TURKISH |
VIROLOGY
- CHAPTER SEVEN
PART TEN
HUMAN
IMMUNODEFICIENCY VIRUS AND AIDS
LOSS OF CD4 CELLS
Dr Richard Hunt
Professor
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
|
|
En
Español |
|
SHQIP - ALBANIAN |
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|

 |
|
THIS CHAPTER IS
IN SEVERAL PARTS
USE THE NEXT>> BUTTON ABOVE TO GO TO THE NEXT PART
TO CONTINUE TO VIROLOGY CHAPTER EIGHT CLICK HERE
|
|
LINKS TO
OTHER HIV AND AIDS SECTIONS ARE AT THE BOTTOM OF THIS PAGE |

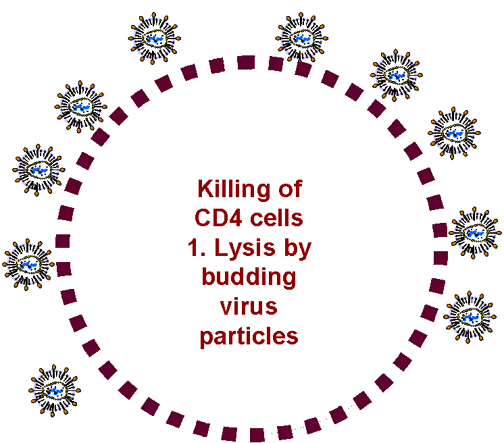
Budding causes cell lysis
 Syncytia formation
Syncytia formation
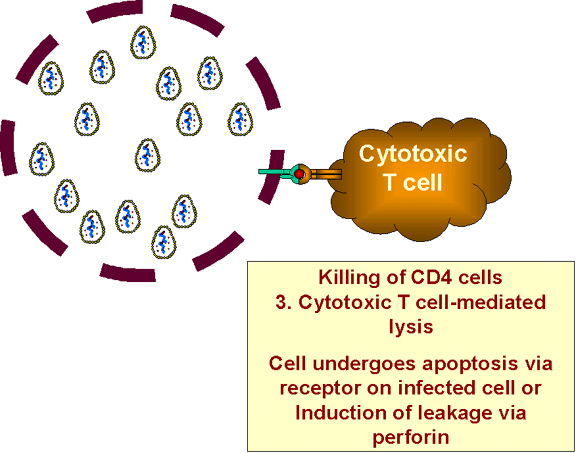
 Infected cells are destroyed by cytotoxic T cells
Infected cells are destroyed by cytotoxic T cells
Figure 27 -
Some possible mechanism for the loss of T4 cells after HIV infection
|
WHY IS
THERE A PROGRESSIVE LOSS OF CD4+ HELPER T CELLS?
WHY DO CD8+ KILLER T CELLS
DISAPPEAR IN THE LATER STAGES OF THE DISEASE?
Why, when only 1 in 10,000 (early) or 1 in 40
(later) cells show productive infection, do all of the T4 cells disappear? It is still
unclear why the CD4+ cells all disappear but there are a number of
possibilities:
-
In an activated, infected CD4 cell, huge numbers of virions are synthesized.
These bud from the cell and result in punctured membranes (figure 27). But the cell needs
to be infected for this to happen and most CD4 cells are not infected.
-
Since the membrane of HIV fuses with the membrane of the cell to be infected
by a pH-independent mechanism, syncytia formation can occur leading to the
spread of virus to uninfected cells (figure 27). But syncytia are not very common.
-
Infected cells that are producing viral proteins (but not those in the latent
state) will present those proteins on the cell surface in association with
class I MHC histocompatibility antigens. The infected cell, like other
virally-infected cells, will be destroyed by cytotoxic T cells (figure 27). Again this only
happens in cells that are infected by HIV.
-
Gp120 is linked to the Gp41 on the
virus surface by non-covalent interactions and is frequently shed from infected
cells or from virus particles. This binds to uninfected cells via
CD4 antigen. As a result, they appear to be infected and are destroyed by the immune
system.
-
There have been reports of AIDS-related cytotoxic antibodies
in infected patients that may react
with a specific antigen on the surface of activated but uninfected T4 cells.
|
| |
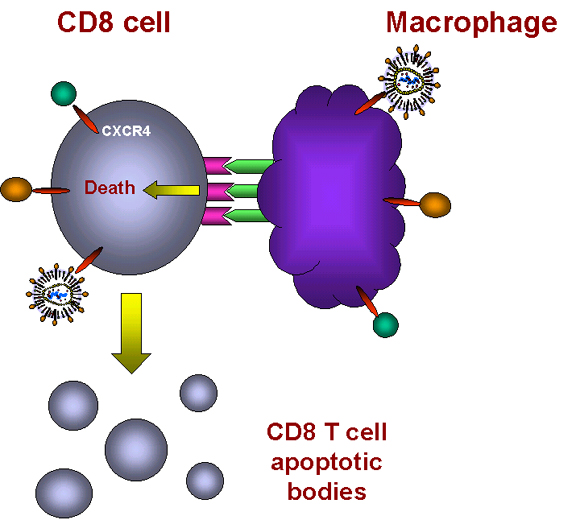
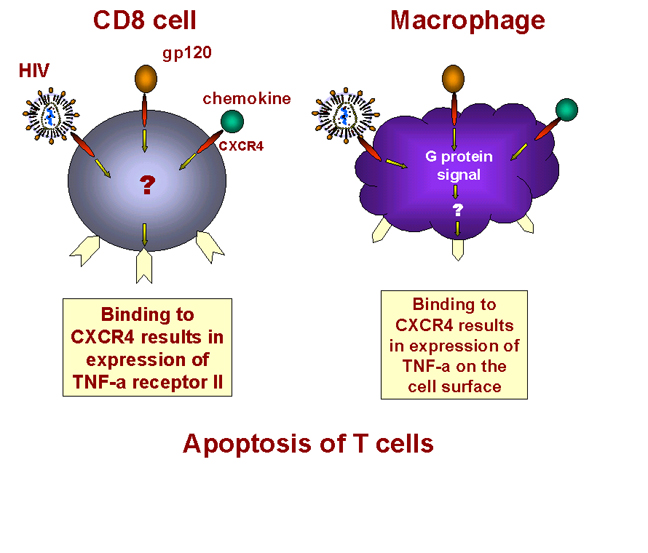
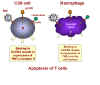 Binding of cytokine induces TNF alpha expression in macrophage and
receptor expression in CD8+ T cell
Binding of cytokine induces TNF alpha expression in macrophage and
receptor expression in CD8+ T cell
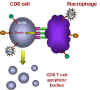 The cells contact one-another and TNF-alpha and the receptor interact.
Apoptosis ensues
The cells contact one-another and TNF-alpha and the receptor interact.
Apoptosis ensues
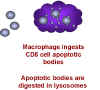 Macrophages internalize T cell
Macrophages internalize T cell
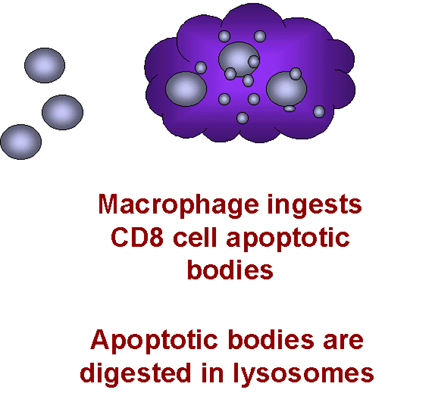
Figure 28 -
Induction of apoptosis in T8 cells
|
-
AIDS may have an auto-immune component. In
a normal antigenic response carried out by T4 cells, CD4 antigen interacts with MHC type II histocompatibility
antigens. Since Gp120 also binds to CD4, the Gp120 can mimic MHC class II
antigens since both have a CD4-binding site; indeed, there appear to
be regions of similar sequence in the two proteins. Thus anti-Gp120 antibodies may turn out to be anti-MHC
antibodies as well. (This might spell trouble for vaccine production).
-
It is possible that HIV might
infect a subset of T4 cells that is vital to propagation of
entire population of T4 cells
-
HIV proteins may alter T4 cell function.
There is some evidence for this.
-
Presently, the most actively
studied possibility for the loss of the entire CD4+ and CD8+ cell population is
that HIV initiates apoptosis in these cells (Such apoptosis is a normal process in
T4 cells to overcome autoimmunity and to terminate an immune response) (figure
28). This is now
thought to be a major factor in the loss of CD4 cells during the progression of
the disease
Some of the above may explain why only a
minority of T4 cells appear to be infected at a given time yet all disappear in the later
stages of the disease. It could also be that the virus switches from one T4 cell
population to another as it switches its co-receptor (see above).
CD8+ cells are only infected by HIV
in small numbers and their levels remain high during the course of the
disease for many years. And then, until recently inexplicably, they rapidly die off.
It appears that some of
the HIV subtypes that occur late in infection prompt a mass apoptosis of CD8 cells.
Although CD8 cells are mostly CD4-, they do have CXCR4 co-receptor and HIV can
bind to this (only the later syncytium-inducing strains of HIV do this). Since
little CD4 antigen is
present there is no infection but binding to CXCR4 sends a signal to the cell, the
signal for apoptosis and mass CD8+ cell suicide ensues.
|
|
|
How does this happen? It
is now known that binding of strains of HIV that arise later in infection to the
CXCR4 receptor sets in motion the tumor necrosis-alpha death transduction pathway
(figure 28). In macrophages, binding of a ligand to CXCR4 receptor on the cell surface
induces the expression of TNF-alpha. In CD8+ T cells, the same binding triggers
the expression of TNF-alpha receptor II.
When such a macrophage and a CD8+ T cell come in contact, the TNF-alpha on the
macrophage binds to the TNF-alpha receptor on the CD8+ T cell. This triggers an
apoptosis signal in the CD8+ T cell resulting in the vesiculation of the CD8+ T
cell (figure 28). Macrophages then phagocytose the remains of the T cell. This explains why
macrophages have to be present for the CD8+ cells to die. Why would this happen
naturally? Why do chemokines act as death signals for CD8+ T cells? These cells
are killer cells and may cause serious trouble if they end up in the wrong
place. It is thought that chemokines direct CD8+ T cells to the fate of
macrophage-mediated death unless they reach their appropriate location.
|
|
|
|
|
|
OTHER SECTIONS ON HIV
PART I HUMAN
IMMUNODEFICIENCY VIRUS AND AIDS
PART II HIV AND AIDS, THE
DISEASE
PART III COURSE OF THE DISEASE
PART IV PROGRESSION AND
COFACTORS
PART V STATISTICS
PART VI SUBTYPES AND
CO-RECEPTORS
PART VII COMPONENTS AND LIFE
CYCLE OF HIV
PART VIII LATENCY OF HIV
PART IX GENOME OF HIV
PART X LOSS OF CD4 CELLS
PART XI POPULATION
POLYMORPHISM
APPENDIX I ANTI-HIV VACCINES
APPENDIX II DOES HIV CAUSE
AIDS?
APPENDIX III ANTI-HIV
CHEMOTHERAPY
|
| |
|
| |
 Return to the Virology Section of Microbiology and Immunology On-line
Return to the Virology Section of Microbiology and Immunology On-line
Return to the
front page of Microbiology and Immunology On-line
This page last changed on
Sunday, August 28, 2016
Page maintained by
Richard Hunt
|
 Binding of cytokine induces TNF alpha expression in macrophage and
receptor expression in CD8+ T cell
Binding of cytokine induces TNF alpha expression in macrophage and
receptor expression in CD8+ T cell