|
x |
x |
|
 |
 |
|
BỆNH NHIỄM TRÙNG |
VI KHUẨN HỌC |
MIỄN DỊCH HỌC |
NẤM HỌC |
KÝ SINH TRÙNG HỌC |
VIRÚT HỌC |
|
ENGLISH
|
MIỄN DỊCH HỌC - CHƯƠNG MƯỜI HAI
ĐÁP ỨNG MIỄN DỊCH QUA TRUNG GIAN TẾ BÀO
Tương tác tế bào-tế bào trong đáp ứng miễn dịch đặc hiệu
Dr Gene Mayer
Professor
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
and
Dr Jennifer Nyland
Assistant Professor
Department of Pathology, Microbiology and Immunology
University of South Carolina School of Medicine
Biên dịch: Nguyễn Văn Đô, MD., PhD.,
Bộ môn Sinh lý bệnh-Miễn dịch,
Trường Đại học Y Hà Nội,
Hà Nội, Việt Nam
|
|
TURKISH |
|
FRANCAIS |
|
PORTUGUES |
|
Let us know what you think
FEEDBACK |
|
SEARCH |
|
|
|

 |
|
Logo image © Jeffrey
Nelson, Rush University, Chicago, Illinois and
The MicrobeLibrary |
|
|
|
Edited and
illustrated by Dr Richard Hunt
|
|
|
|
MỤC TIÊU GIẢNG DẠY
Thảo luận về vai trò trung tâm của các tế bào Th trong
các đáp ứng miễn dịch
Mô tả sự tương tác tế bào-tế bào diễn ra trong (i) đáp ứng kháng thể với
kháng nguyên phụ thuộc T, (ii) hình thành các tế bào T gây độc tế bào,
và (iii) hoạt hóa các đại thực bào và tế bào NK
Thảo luận về cơ chế làm chết tế bào bởi các tế bào T độc và các tế bào
NK
Thảo luận các đáp ứng với kháng nguyên không phụ thuộc
T
|
VAI TRÒ TRUNG TÂM CỦA TẾ BÀO Th
TRONG ĐÁP ỨNG MIỄN DỊCH
Như mô tả trong Hình 1, sau khi các tế bào
Th nhận ra kháng nguyên đặc hiệu được trình diện bởi một tế bào trình diện kháng
nguyên (APC), chúng khởi động một số quá trình miễn dịch quan trọng, bao gồm:
-
Lựa chọn cơ chế hiệu ứng thích hợp (ví
dụ, hoạt hóa tế bào B, hoặc tạo ra Tc);
-
Cảm ứng để phát triển thành các tế bào
phù hợp
-
Tăng cường các hoạt động chức năng của
các tế bào khác (ví dụ, bạch cầu hạt, đại thực bào, tế bào NK ).
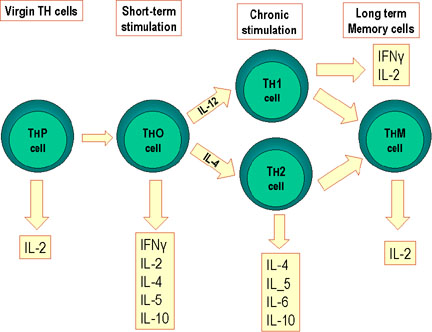
Có bốn tiểu quần thể các tế bào Th: Th0, Th1, Th2 và Th17. Khi các tế bào Th0
trinh tiết gặp phải kháng nguyên trong các mô bạch huyết thứ phát, chúng có khả
năng biệt hóa thành các tế bào Th1 trong viêm, các tế bào Th2 trợ giúp hoặc tế
bào Th17 gây bệnh, được phân biệt bởi các cytokin mà chúng sản xuất ra (Hình 2).
Một số tế bào Th0 trở thành Th1, Th2 hoặc T17 phụ thuộc vào các cytokin trong
môi trường, đó là ảnh hưởng bởi kháng nguyên. Ví dụ một số kháng nguyên kích
thích sản xuất IL-4 có ưu thế tạo ra các tế bào Th2, trong khi kháng nguyên khác
kích thích sản xuất IL-12, ưu tiên tạo ra các tế bào Th1. Tế bào Th1, Th2 và
Th17 ảnh hưởng đến các tế bào khác nhau cũng như các loại đáp ứng miễn dịch, như
trong Hình 3 minh họa cho các tế bào Th1 và Th2.
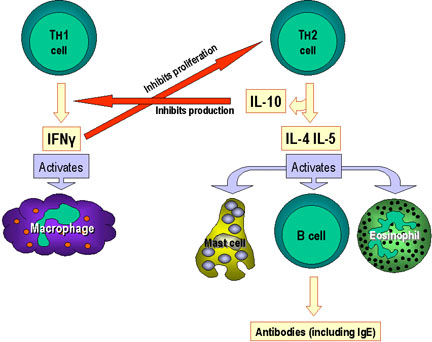
Cytokin được sản xuất bởi Th1 sẽ làm hoạt
hóa các đại thực bào và tham gia vào việc tạo ra các tế bào lympho T gây độc (CTL),
dẫn đến đáp ứng miễn dịch qua trung gian tế bào. Ngược lại, cytokin được sản
xuất bởi Th2 sẽ giúp đỡ để hoạt các tế bào B, kết quả là sản xuất ra kháng thể.
Trong một phát hiện gần đây, các tế bào Th17
(được gọi là như vậy vì nó sản xuất IL-17) biệt hóa (ở người) để đáp ứng với
IL-1, IL-6, và IL-23. TGF-β là chất quan trọng đối với sự biệt hóa Th17 ở chuột,
nhưng không phải ở người. IL-17 làm nặng thêm một số bệnh tự miễn dịch bao gồm
cả bệnh đa xơ cứng, bệnh viêm ruột, và viêm khớp dạng thấp. IFN-γ được sản xuất
bởi các tế bào Th1 ức chế sự phát triển các tế bào Th2 và sự biệt hóa của các tế
bào Th17; IL-10 được sản xuất bởi các tế bào Th2 ức chế sự sản xuất IFN-γ bởi
các tế bào Th1. Ngoài ra, mặc dù không được chỉ ra, IL-4 ức chế sự sản xuất các
tế bào Th1 và sự biệt hóa của các tế bào Th17. Vì vậy, các đáp ứng miễn dịch
được hướng đến loại đáp ứng cần thiết để đối phó với tác nhân gây bệnh gặp phải
- đáp ứng miễn dịch qua trung gian tế bào dùng cho các tác nhân gây bệnh nội bào
hoặc đáp ứng kháng thể đối với tác nhân gây bệnh ngoại bào.
|
|
TỪ KHÓA
Tế bào Th1
Tế bào Th2
Mô hình chất mang-hapten
CD28
B7
CD40
Phối tử CD40
CD5
Tế bào B1
B2
CTL
Phối tử Fas
Perforin
Granzym
Caspase
IFN-γ
Hoạt hóa
|
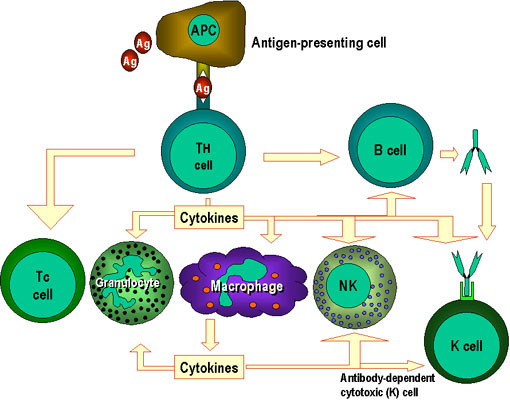
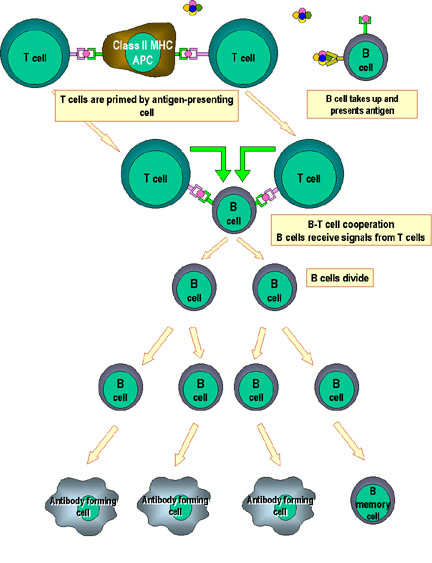
 Hình 1
Hình 1
Các tế bào Th là trung tâm của miễn dịch qua trung gian tế bào. Các tế bào
trình diện kháng nguyên trình kháng nguyên cho các tế bào T hỗ trợ (Th). Tế
bào Th nhận biết các epitop đặc hiệu, được chọn là epitop đích. Các cơ chế
hiệu ứng phù hợp hiện nay đã được xác định. Ví dụ, các tế bào Th giúp các tế
bào B để sản xuất kháng thể và cũng có thể hoạt hóa các tế bào khác. Các tín
hiệu hoạt hóa được sản xuất bởi các tế bào Th là cytokin (các lymphokin)
nhưng cytokin tương tự được sản xuất bởi các đại thực bào và tế bào khác
cũng tham gia vào quá trình này.
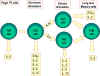 Hình 2
Hình 2
Sự biệt hóa các tế bào Th của chuột. Các tế bào Th chuột biệt hóa thành các
dưới nhóm để tổng hợp các loại lymphokin khác nhau. Điều này cũng xảy ra ở
người.
 Hình 3
Hình 3
Lựa chọn cơ chế hiệu ứng bởi các tế bào Th1 và Th2. Ngoài việc xác định các
con đường hiệu ứng khác nhau nhờ sản xuất lymphokin của chúng, các tế bào
Th1 kiểm soát tế bào Th2 và ngược lại
|
|
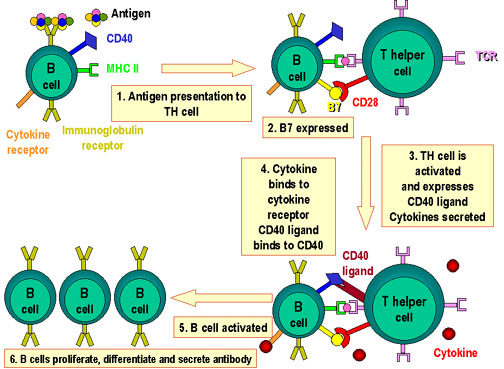
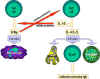
Hình 4
Các phân
tử liên quan đến sự tương tác của các tế bào B và Th.
Kháng
 nguyên được xử lý bởi tế bào B. Đồng kích thích được thể
hiện.
nguyên được xử lý bởi tế bào B. Đồng kích thích được thể
hiện.
 Các peptid kháng nguyên được trình diện trong
phân tử MHC lớp II. Các tế bào T nhận ra peptid cùng với các kháng
nguyên MHC và các đồng kích thích. Các tế bào T biểu lộ phối tử CD40 sau
đó liên kết với kháng nguyên CD40 trên tế bào B và tế bào B phân chia và
biệt hóa. Kháng thể được sản xuất bởi các tế bào B
Các peptid kháng nguyên được trình diện trong
phân tử MHC lớp II. Các tế bào T nhận ra peptid cùng với các kháng
nguyên MHC và các đồng kích thích. Các tế bào T biểu lộ phối tử CD40 sau
đó liên kết với kháng nguyên CD40 trên tế bào B và tế bào B phân chia và
biệt hóa. Kháng thể được sản xuất bởi các tế bào B
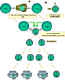 Hình 5
Hình 5
Sự hợp tác của các tế bào trong phản ứng miễn dịch.
Các tế bào trình diện kháng nguyên (ví dụ như tế bào đuôi gai) trình
diện kháng nguyên đã được xử lý cho tế bào T nghỉ, do đó nó sẽ mồi chúng.
Các tế bào B cũng xử lý kháng nguyên và trình diện cho tế bào T. Sau đó,
chúng nhận được tín hiệu từ các tế bào T và trở nên phân chia và biệt
hóa. Một số tế bào B trở thành các tế bào tạo kháng thể và một số ít tế
bào trở thành tế bào B nhớ.
|
TƯƠNG TÁC TẾ BÀO-TẾ BÀO TRONG ĐÁP ỨNG
SINH KHÁNG THỂ ĐỐI VỚI KHÁNG NGUYÊN NGOẠI SINH PHỤ THUỘC T
Mô hình chất mang-hapten
Trong lịch sử một trong những phát hiện quan
trọng của miễn dịch học là cả hai loại tế bào T và B được yêu cầu để sản
xuất kháng thể cho một loại phức hợp protein. Một đóng góp lớn cho sự hiểu
biết của chúng ta về quá trình này là từ các nghiên cứu về sự hình thành các
kháng thể chống hapten. Nghiên cứu về chất mang-hapten được hình thành là:
-
Tế bào Th2 nhận biết các quyết định kháng
nguyên của chất mang và các tế bào B nhận biết các quyết định của hapten
-
Tương tác giữa các tế bào B đặc hiệu với
hapten và các tế bào Th đặc hiệu chất mang được giới hạn trong MHC tự
thân
-
Tế bào B có cả hai chức năng nhận biết và
trình diện kháng nguyên
Các tế bào B chiếm một vị trí độc tôn trong các đáp ứng miễn dịch vì chúng
có cả hai chức năng là tiền thân tế bào sản xuất ra kháng thể và biểu lộ các
phân tử MHC lớp II trên bề mặt. Do đó, chúng có khả năng sản xuất kháng thể
có độ đặc hiệu giống như kháng thể có vai trò thụ thể của chúng. Ngoài ra,
chúng có chức năng như một tế bào trình diện kháng nguyên. Trong phạm vi của
mô hình liên hợp chất mang-Hapten, cơ chế được thể hiện như sau: hapten này
được nhận biết bởi các kháng thể làm thụ thể, chất mang-hapten được đưa vào
tế bào B, xử lý, và các mảnh peptid của protein của chất mang được trình
diện cho tế bào T hỗ trợ. Hoạt hóa của các tế bào T làm sản xuất các cytokin
cho phép các tế bào B đặc hiệu hapten trở nên hoạt hóa để sản xuất kháng thể
hòa tan chống hapten. Hình 4 tóm tắt sự tương tác của các tế bào B-T.
Lưu ý rằng có nhiều tín hiệu gửi đến các tế
bào B trong mô hình tương tác giữa các tế bào Th2 và tế bào B. Như trường
hợp để hoạt hóa các tế bào T, các tín hiệu thu được từ TCR nhận biết một
phân tử MHC-peptid chưa đủ để hoạt hóa tế bào T, do đó tế bào B cũng vậy. Sự
kết hợp của một kháng nguyên vào các kháng thể làm thụ thể sẽ cung cấp một
tín hiệu cho tế bào B, nhưng tín hiệu đó chưa đủ. Tín hiệu thứ hai được tạo
ra bởi các phân tử đồng kích thích là cần thiết, phân tử quan trọng nhất
trong số này là CD40L trên tế bào T có liên kết với CD40 trên tế bào B để
bắt đầu chuyển tín hiệu thứ hai.
Tương tác tế bào-tế bào trong đáp ứng sinh kháng
thể lần thứ nhất
Các tế bào B không phải là tế bào trình trình
diện kháng nguyên tốt nhất trong đáp ứng sinh kháng thể lần thứ nhất; tế bào
đuôi gai hay đại thực bào có hiệu quả hơn. Tuy nhiên, với một số thay đổi
nhỏ ở mô hình chất mang-hapten của các tương tác tế bào-tế bào mô tả ở trên
cũng áp dụng cho sự tương tác tế bào-tế bào trong đáp ứng kháng thể lần thứ
nhất (Hình 5). Trong đáp ứng lần đầu, trước tiên các tế bào Th2 gặp kháng
nguyên được trình diện bởi các tế bào đuôi gai hay đại thực bào. Các tế bào
Th2 "mồi" sau đó có thể tương tác với các tế bào B đã bắt kháng nguyên và
đang trình diện kháng nguyên peptid gắn với các phân tử MHC lớp II. Các tế
bào B cần có hai tín hiệu để hoạt hóa - một là sự liên kết của kháng nguyên
với các kháng thể bề mặt và tín hiệu thứ hai được xuất phát từ sự tham gia
của phối tử CD40/CD40 trong khi tương tác tế bào-tế bào B/Th2. Ngoài ra, các
cytokin được sản xuất bởi các tế bào Th2 giúp các tế bào B phân chia và biệt
hóa thành các tế bào tương bào sản xuất ra kháng thể.
Tương tác tế bào-tế bào trong đáp
ứng kháng thể lần thứ hai
Sau đáp ứng miễn dịch lần tứ nhất, nhiều tế
bào T và B trí nhớ được hình thành. Các tế bào B nhớ có thụ thể là kháng thể
bề mặt có ái tính cao (do trưởng thành của ái tính), cho phép chúng kết hợp
và trình diện kháng nguyên ở nồng độ thấp hơn nhiều so với yêu cầu đối với
đại thực bào hoặc tế bào đuôi gai. Ngoài ra, các tế bào T nhớ được hoạt hóa
dễ dàng hơn so với các tế bào T trinh tiết. Vì vậy, sự tương tác B/Th là đủ
để tạo ra đáp ứng kháng thể thứ cấp. Nó không cần thiết phải mồi các tế bào
T nhớ (mặc dù nó có thể xảy ra) với các tế bào trình diện kháng nguyên bởi
các tế bào đuôi gai hay đại thực bào.
Cytokin và chuyển lớp kháng thể
Cytokin được sản xuất bởi các tế bào Th2
không chỉ hoạt hóa sự phát triển và biệt hóa của các tế bào B, chúng cũng
giúp điều hòa sản xuất các lớp kháng thể. Các cytokin khác nhau ảnh hưởng
đến việc chuyển lớp khác nhau của các kháng thể có chức năng khác nhau. Bằng
cách này, đáp ứng kháng thể được thay đổi để phù hợp với các tác nhân gây
bệnh gặp phải (ví dụ như sản xuất các kháng thể IgE trong nhiễm giun ký sinh).
Bảng 1 cho thấy tác dụng của cytokin khác nhau trên các loại kháng thể được
sản xuất.
|
Cytokin
|
IgG1
|
IgG2a
|
IgG2b
|
IgG3
|
IgA
|
IgE
|
IgM
|
|
IL-4
|
Cảm ứng
|
Ức chế
|
|
Ức chế
|
|
Cảm ứng
|
Ức chế
|
|
IL-5
|
|
|
|
|
Tăng sản xuất
|
|
|
|
IFN-gamma
|
Ức chế
|
Cảm ứng
|
|
Cảm ứng
|
|
Ức chế
|
Ức chế
|
|
TGF-beta
|
|
|
Cảm ứng
|
Ức chế
|
Cảm ứng
|
|
Ức chế
|
|
Điều hòa Isotyp bởi cytokin của tế
bào T chuột
Một số cytokin gây ra (màu xanh lá cây) hoặc ức chế (màu hồng) sản xuất
một số loại kháng thể nhất định. Sự ức chế là do chuyển đổi các isotyp
khác nhau
|
|
Bảng 1 |
|
| |
TƯƠNG TÁC TẾ BÀO-TẾ BÀO TRONG CÁC ĐÁP ỨNG VỚI KHÁNG NGUYÊN NGOẠI SINH KHÔNG PHỤ
THUỘC T
Các đáp ứng
sinh kháng thể với kháng nguyên không phụ thuộc T không cần tương tác tế bào-tế
bào. Bản chất cao phân tử của các kháng nguyên này cho phép liên kết chéo của
các thụ thể kháng nguyên trên tế bào B dẫn đến hoạt hóa. Không có đáp ứng lần
thứ hai, ái tính trưởng thành hay chuyển lớp kháng thể. Đáp ứng với kháng nguyên
không phụ tuộc T là do sự hoạt hóa của một tiểu quần thể các tế bào B được gọi
là tế bào B CD5+ (còn gọi là tế bào B1), chúng khác với các tế bào B thông
thường có CD5-(còn gọi là tế bào B2).
Tế bào CD5+ (B1)
Các tế
bào CD5+ là những tế bào B đầu tiên xuất hiện trong sự phát triển cá thể.
Chúng biểu lộ IgM bề mặt, nhưng có ít hoặc không có IgD và chủ yếu chúng sản
xuất các kháng thể IgM từ gen dòng mầm đột biến soma tối thiểu. Các kháng
thể được sản xuất bởi các tế bào này có ái lực thấp và thường đa phản ứng (kết
hợp nhiều kháng nguyên). Hầu hết IgM trong huyết thanh có nguồn gốc từ các
tế bào B CD5+. Tế bào B CD5+ không phát triển thành các tế bào trí nhớ. Một
đặc tính quan trọng của các tế bào này là chúng được tự đổi mới, không giống
như các tế bào B thông thường là phải được thay thế từ tủy xương. Các tế bào
B CD5+ được tìm thấy trong các mô ngoại vi và là tế bào B chiếm ưu thế trong
khoang phúc mạc. Các tế bào B1 là hàng rào chủ yếu chống lại nhiều tác nhân
gây bệnh là vi khuẩn có polysaccharid ở thành tế bào. Tầm quan trọng của các
tế bào này trong hệ miễn dịch được chỉ ra bởi thực tế là nhiều cá thể có
khiếm khuyết tế bào T vẫn có thể chống lại các tác nhân gây bệnh là vi khuẩn.
|
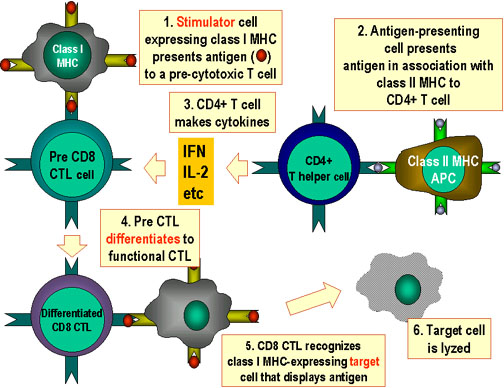
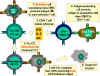 Hình
6 Hình
6
Tế bào CTL phải phân chia trong đáp ứng với kháng nguyên. Để phân chia
thành các tế bào có chức năng gây độc tế bào, tế bào tiền lympho T gây
độc có CD8+ phải nhận được hai tín hiệu khác nhau. Đầu tiên, chúng phải
nhận ra kháng nguyên trình diện trên phân tử MHC-I (các tế bào kích
thích) và, thứ hai, chúng phải được kích thích bởi cytokin. IL-2,
interferon gamma-và những chất khác được tạo ra bởi các tế bào T CD4+ là
kết quả của sự tương tác của chúng với kháng nguyên trình diện trên MHC-II.
Với kết quả của hai tín hiệu, các tiền CTL biệt hóa thành CTL hoạt hóa
và có thể ly giải tế bào đích có cùng kháng nguyên.
Adapted from Abbas, et. al. Cellular and Molecular Immunology.
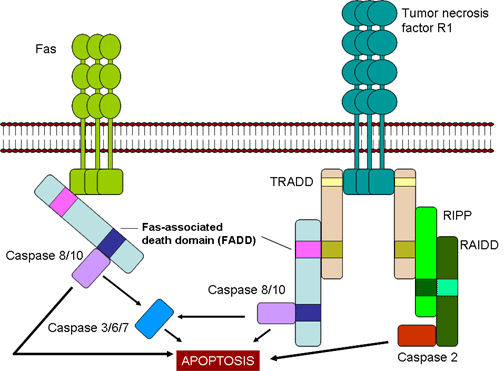
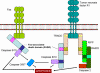 Hình 7
Hình 7
Các tế bào CTL giết tế bào đích thông qua Fas-và TNF
|
TƯƠNG TÁC TẾ BÀO-TẾ BÀO TRONG ĐÁP ỨNG MIỄN DỊCH TẾ BÀO-
SỰ HÌNH THÀNH TẾ BÀO Tc TRONG ĐÁP ỨNG VỚI CÁC KHÁNG NGUYÊN NỘI SINH Ở BÀO TƯƠNG
Các tế bào lympho T gây độc tế bào chưa hoàn toàn trưởng thành
khi chúng rời khỏi tuyến ức. Chúng có một TCR có chức năng nhận biết kháng
nguyên, nhưng chúng không thể ly giải một tế bào đích. Chúng phải biệt hóa thành
các tế bào Tc có đầy đủ các chức năng hiệu ứng. Các tế bào gây độc biệt hóa từ
một "tiền CTL" để đáp ứng với hai tín hiệu:
- Có kháng nguyên đặc hiệu trên phân tử MHC lớp I nằm trên
một tế bào kích thích
- Cytokin sản xuất bởi tế bàoTh1, đặc biệt là IL-2 và IFN-gamma.
Điều này được thể hiện trong hình 6.
Các đặc tính của ly giải qua trung gian CTL
- Tiêu diệt bởi CTL là đặc hiệu kháng nguyên. Để bị
tiêu diệt bởi một CTL, các tế bào đích phải mang kháng nguyên giống với
kháng nguyên trên MHC lớp I mà đã hoạt hóa sự biệt hóa tiền CTL.
- Sự tiêu diệt bởi CTL cần phải có sự tiếp xúc tế bào.
CTL được hoạt hóa để giết khi chúng nhận ra những kháng nguyên đích liên
kết với phân tử MHC bề mặt tế bào. Các tế bào lân cận không có các
MHC-kháng nguyên phù hợp sẽ không bị ảnh hưởng.
- Các CTL không bị tổn thương khi chúng ly giải tế bào
đích. Mỗi CTL có khả năng giết chết nhiều tế bào đích một cách liên tục
Các cơ chế tiêu diệt qua trung gian CTL
Các CTL sử dụng một số cơ chế để tiêu diệt tế bào đích,
một số CTL trong số đó phải tiếp xúc trực tiếp tế bào-tế bào và những CTL
khác thì nhờ vào việc sản xuất các cytokin. Trong tất cả các trường hợp sự
chết của tế bào đích là kết quả của sự chết theo chương trình.
- Tiêu diệt qua trung gian Fas và TNF (Hình 7)
Sau khi được tạo ra, các CTL biểu lộ phối tử Fas trên bề mặt của chúng
và các phối tử Fas sẽ liên kết với các thụ thể Fas trên các tế bào đích.
Ngoài ra, TNF-α được tiết ra bởi CTLs có thể gắn vào các thụ thể TNF
trên các tế bào đích. Các thụ thể Fas và TNF là một gia đình các thụ thể
liên quan chặt chẽ với nhau, khi chúng gặp phải phối tử của chúng làm
thành các thụ thể bộ ba. Các thụ thể này cũng có vùng chết nằm trong bào
tương mà sau khi bộ ba hóa chúng có thể kích hoạt các caspase để gây ra
chết theo chương tình của tế bào đích.
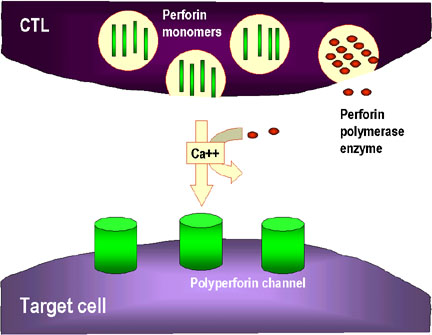
- Tiêu diệt bởi hạt chứa enzym (Hình 8)
Các CTL hoàn chỉnh có nhiều hạt nhỏ có chứa perforin và granzyme. Khi
tiếp xúc với tế bào đích, perforin được giải phóng và nó trùng hợp để
hình thành các kênh trong màng tế bào đích. Granzyme, đó là protease
serine được đưa vào các tế bào đích thông qua các kênh này và hoạt hóa
caspase và nuclease trong các tế bào đích dẫn đến chết theo chương trình.
|
| |
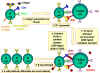
Hình 8
Cơ chế phá hủy các tế bào đích bởi CTL
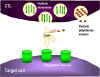 1. CTL làm vỡ hạt và giải phóng perforin đơn phân vào môi trường xung quanh. Các
enzym trùng hợp perforin để hình thành các kênh polyperforin cũng được hình
thành và cùng với ion Ca++ xúc tác hình thành các kênh thủy phân trong màng của
tế bào đích.
1. CTL làm vỡ hạt và giải phóng perforin đơn phân vào môi trường xung quanh. Các
enzym trùng hợp perforin để hình thành các kênh polyperforin cũng được hình
thành và cùng với ion Ca++ xúc tác hình thành các kênh thủy phân trong màng của
tế bào đích.
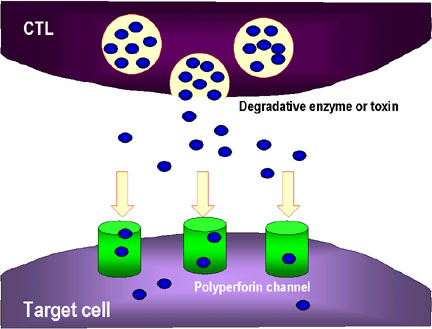
 2. CTL cũng có thể giải phóng ra các enzym phân hủy và các chất độc,
chúng đi qua các kênh perforin và làm tổn thương các tế bào đích.
2. CTL cũng có thể giải phóng ra các enzym phân hủy và các chất độc,
chúng đi qua các kênh perforin và làm tổn thương các tế bào đích.
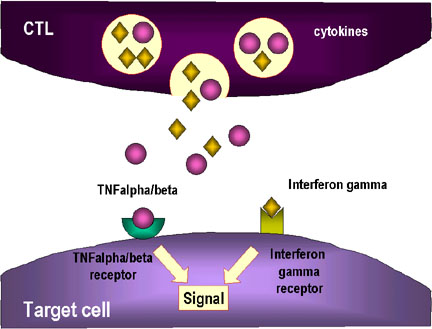
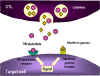 3. Các cytokin như TNF alpha và TNF beta được sản xuất bởi các CTL hoặc
đại thực bào ở gần đó. Interferon gamma cũng có thể được tạo ra từ các
CTL hoặc từ các tế bào bạch huyết gần đó. Chúng liên kết với các thụ thể
trên các tế bào đích và gây ra chết theo chương trình.
3. Các cytokin như TNF alpha và TNF beta được sản xuất bởi các CTL hoặc
đại thực bào ở gần đó. Interferon gamma cũng có thể được tạo ra từ các
CTL hoặc từ các tế bào bạch huyết gần đó. Chúng liên kết với các thụ thể
trên các tế bào đích và gây ra chết theo chương trình. |
|
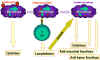 Hình 9
Hình 9
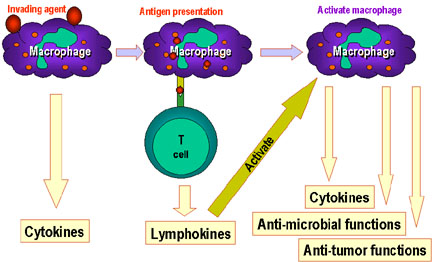
Các đại thực bào đóng vai trò quan trọng của hệ thống miễn dịch. Trước
khi các tế bào miễn dịch B và T bắt đầu thực hiện chức năng. Các đại
thực bào trình diện kháng nguyên cho tế bào T, sau đó tế bào T sẽ tiết
ra lymphokin hoạt hóa đại thực bào để thực hiện nhiều chức năng khác bao
gồm cả việc sản xuất thêm cytokin
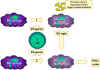 Hình 10
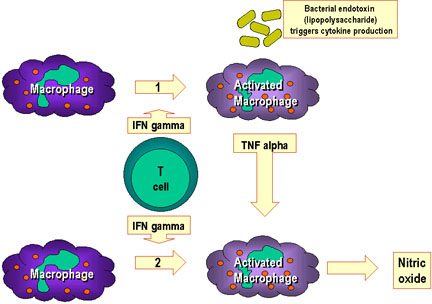
Hình 10
Đại thực bào được hoạt hóa nhờ sự tương tác của nhiều cytokin và các yếu
tố khác.
Trong con đường 1, TNF alpha-được tiết ra từ đại thực bào là kết quả của
sự tương tác của interferon gamma với các thành phần của vi khuẩn gây ra
sản xuất cytokin. Ví dụ thành phần lipopolysaccharid. Các TNF-alpha từ
con đường 1 làm tăng tiết oxit nitric bởi đại thực bào hoạt hóa do
interferon trong con đường thứ 2.
|
TƯƠNG TÁC TẾ BÀO-TẾ BÀO
TRONG ĐÁP ỨNG MIỄN DỊCH TẾ BÀO- SỰ HOẠT HÓA ĐẠI THỰC BÀO TRONG ĐÁP ỨNG
VỚI CÁC KHÁNG NGUYÊN NỘI SINH
Các đại thực bào đóng vai trò trung
tâm trong hệ thống miễn dịch. Như trong hình 9, các đại thực bào có liên
quan đến:
-
Hàng rào đầu tiên, là một phần
của hệ miễn dịch tự nhiên
-
Trình diện kháng nguyên cho các
tế bào Th
-
Có nhiều chức năng hiệu ứng khác
nhau (ví dụ, sản xuất cytokin, hoạt tính diệt khuẩn và khối u).
Thực tế là các đại thực bào đóng vai
trò quan trọng không chỉ ở khả năng miễn dịch mà còn trong tái tạo mô.
Tuy nhiên, do khả năng hoạt động của chúng, đại thực bào cũng có thể làm
tổn hại đến các mô. Bảng 2 tóm tắt các chức năng của đại thực bào trong
miễn dịch và viêm nhiễm.
Viêm-Sốt
Sản xuất:
IL-6, TNF alpha, IL-1 – là chất gây sốt
|
Tổn thương mô
Enzym thủy phân
Sản xuất oxy già
C3a của bổ thể
Sản xuất TNF alpha
|
Miễn dịch
Lympho hoạt hóa tiết ra:
IL-12 được tiết ra do Th1 hoạt hóa
IL-10 được tiết ra do Th2 hoạt hóa
Hoạt hóa lympho:
Tiết ra IL-1
Xử lý và trình diện kháng nguyên
|
Hoạt tính kháng khuẩn
Các sản phẩm phụ thuộc oxy:
Oxy già
Chất oxy hóa
Gốc tự do
Acid hypochlorit
Các sản phẩm không phụ
thuộc oxy
Thủy phân môi trường acid
Protein cation
lysozyme
|
Tái tạo mô
Tiết ra một số yếu tố:
Enzym phân cắt (elastase, hyaluronidase,collagenase)
Yế tố kích thích tế bào xơ non
Kích thích tạo mạch
|
Hoạt tính chống ung thư
Yếu tố gây độc
Oxy già
C3a của bổ thể
Protease
Arginase
Oxid nitric
TNF alpha
|
|
Bảng 2 |
Nhiều chức năng của đại thực bào có thể được thực
hiện bởi các đại thực bào hoạt hóa. Hoạt hóa đại thực bào có thể được
định nghĩa là thay đổi về lượng trong biểu lộ sản phẩm gen khác nhau,
cho phép các đại thực bào hoạt hóa để thực hiện một số chức năng mà
không thể được thực hiện bởi các đại thực bào ở trạng thái nghỉ.
Hoạt hóa đại thực bào là một chức năng quan trọng
của tế bào Th1. Khi các tế bào Th1 được hoạt hóa bởi một APC như đại
thực bào thì chúng giải phóng ra IFN-γ, đó là một trong hai tín hiệu cần
thiết để hoạt hóa một đại thực bào. Lipopolysaccharid (LPS) từ vi khuẩn
hoặc TNF-α được sản xuất bởi các đại thực bào tiếp xúc với các sản phẩm
vi khuẩn sẽ cung cấp tín hiệu thứ hai (Hình 10).
Các cơ chế hiệu ứng sử dụng bởi các đại thực bào,
bao gồm sản xuất:
-
TNF-α, có thể gây ra chết theo chương trình
-
Oxit nitric và các chất trung gian khác chứa
nitơ
-
Phản ứng qua oxy
-
Protein cation và enzym thủy phân
Hoạt hóa đại thực bào bởi tế bào Th1 là rất quan
trọng trong việc bảo vệ chống lại tác nhân gây bệnh khác nhau. Ví dụ,
phế cầu khuẩn, một tác nhân gây bệnh ngoại bào được kiểm soát ở những
người bình thường bởi các đại thực bào hoạt hóa; tuy nhiên, nó là một
nguyên nhân phổ biến gây tử vong ở bệnh nhân AIDS vì họ thiếu các tế bào
Th1. Tương tự như vậy, vi khuẩn lao, một tác nhân gây bệnh nội bào cư
trú trong các túi ở bào tương và không bị giết bởi các đại thực bào một
cách hiệu quả, trừ khi chúng được hoạt hóa; vì vậy, bệnh nhiễm trùng này
là một vấn đề ở những bệnh nhân AIDS.
|
| |
TƯƠNG TÁC TẾ BÀO-TẾ BÀO
TRONG ĐÁP ỨNG MIỄN DỊCH TẾ BÀO-HOẠT HÓA TẾ BÀO NK
Cytokin được sản xuất bởi các tế bào
Th1 hoạt hóa, đặc biệt là Il-2 và IFN-γ, cũng hoạt hóa các tế bào NK để
trở thành các tế bào tiêu diệt (tế bào LAK). Tế bào LAK có khả năng giết
chết tế bào nhiễm virút và tế bào khối u mà không bị giới hạn MHC. Thật
vậy, sự nhạy cảm của các tế bào đích bị tiêu diệt bởi các tế bào NK và
LAK tỉ lệ nghịch với sự biểu lộ các phân tử MHC lớp I (xem bài giảng về
miễn dịch tự nhiên). Các cơ chế hiệu ứng được sử dụng bởi các tế bào NK
và LAK để diệt các tế bào đích là tương tự như được sử dụng bởi CTLs (ví
dụ, perforin và granzyme). Tế bào NK và LAK cũng có thể tiêu diệt các tế
bào đích phủ kháng thể bởi ADCC.
|
|
|
|
|
|
 Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
Trở về phần Miễn dịch của Vi khuẩn học và Miễn dịch học
online
Page maintained by
Richard Hunt
|
 Hình
6
Hình
6