![]()

VIROLOGIE - CHAPITRE UN
VIROLOGIE DE BASE: DÉFINITIONS, STRUCTURE ET CLASSIFICATION
Dr Margaret Hunt
University of South Carolina School of Medicine
Columbia SC
USADr Dorian McIlroy
Université de Nantes
France
FEEDBACK
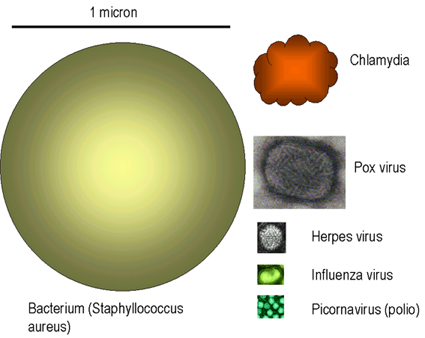 Figure 1
Figure 1
Taille relative de virus et de bactéries Adapted from Koneman et al. Color Atlas and Textbook of
Microbiology 5th Ed. 1997 Virus images © Dr Linda Stannard,
University of Cape Town. Used with permission
Les virus sont constitués d'un acide nucléique (ADN ou ARN) associé à des
protéines codées par l'acide nucléique. Une particule virale peut aussi disposer
d'une membrane en bicouche lipidique (ou enveloppe), qui est acquise à partir de
la cellule hôte, généralement par bourgeonnement à travers une membrane de
cellule hôte. Si une membrane est présente, elle doit contenir une ou plusieurs
glycoprotéines virales qui agissent en tant que ligands pour des récepteurs de
la cellule hôte.
De nombreux virus codent pour une ou plusieurs protéines structurales (celles
qui composent la particule virale mature (ou virion)) et éventuellement une
enzyme qui participe à la réplication du génome viral. Un exemple de virus de ce
type est le circovirus porcin, dont le génome code pour seulement deux protéines:
une protéine qui forme la particule virale, une enzyme qui permet la réplication
de l'ADN génomique du virus. D'autres virus ont des génomes beaucoup plus
complexes, et peuvent coder pour plusieurs dizaines, voire des centaines de
protéines. La plupart de ces protéines ne se retrouvent pas dans le virion
mature, mais participent d'une certaine manière dans la réplication virale. Par
exemple, le cytomégalovirus (CMV) est l'un des virus les plus complexes, et son
génome code pour environ 170 protéines.
Néanmoins, même les virus les plus complexes sont dépendants des enzyme et des
ribosomes de l'hôte pour la production des particules virales. La dépendance des
virus vis-à-vis de la cellule hôte a longtemps rendu difficile le développement
de médicaments antiviraux car la plupart des molécules capables d'inhiber la
réplication virale dans un système de culture cellulaire n'ont pas d'activité
spécifiquement antivirale – ils inhibent des processus cellulaires exploités par
des virus.
Une raison majeure pour étudier la biochimie et la biologie moléculaire des
virus est de trouver des médicaments qui inhibent sélectivement la réplication
virale. Si on peut déterminer à quel moment le virus utilise ses propres
protéines pour une partie de son cycle de réplication, on pourra alors essayer
de développer des médicaments qui inhibent spécifiquement les protéines virales
(et en particulier, les enzymes virales). Les inhibiteurs de protéase utilisés
pour le traitement de l'infection par le VIH sont des exemples de médicaments
conçus et développés après des recherches fondamentales sur le cycle de
réplication viral.
Contrairement aux virus, les bactéries sont beaucoup plus grandes (Figure 1),
effectuent leurs propres processus métaboliques et codent pour toutes les
enzymes dont elles ont besoin. Même lorsque la réaction catalysée est similaire,
les enzymes bactériennes se distinguent de leurs homologues eucaryotes et
peuvent donc être visées par antibiotiques spécifiques. Les tétracyclines, par
exemple, sont des antibiotiques qui ciblent spécifiquement les ribosomes
bactériens, et n'inhibent pas les ribosomes des cellules animales.
Comme les virus, certaines bactéries (comme les mycoplasmes, des Rickettsiae et
des Chlamydiae) peuvent pénétrer dans le cytoplasme des cellules eucaryotes et
devenir des parasites. Néanmoins, ces petites bactéries intracellulaires portent
toutes les enzymes nécessaires à la réplication de leur ADN, et à la synthèse
des protéines. Ainsi, les thérapies antibactériens, y compris ceux qui cible les
bactéries ayant un mode de vie parasitaire, sont plus faciles à développer que
pour les virus.
|
Caractéristiques permettant de distinguer les bactéries des virus |
||||||
|
|
Croissance sur des milieux artificiels ? |
Division par fission binaire |
ARN plus ADN dans la particule infectieuse |
Présence de ribosomes |
Paroi formée de peptidoglycane |
Sensible aux antibiotiques |
|
Bacteria |
Oui |
Oui |
Oui |
Oui |
Oui |
Oui |
|
Mycoplasma |
Oui |
Oui |
Oui |
Oui |
Non |
Oui |
|
Rickettsia |
Non |
Oui |
Oui |
Oui |
Oui |
Oui |
|
Chlamydia |
Non |
Oui |
Oui |
Oui |
Non |
Oui |
|
Viruses |
Non |
Non |
Non |
Non * |
Non |
Non |
|
* Les virus de la famille Arenaviridae semblent incorporer des ribosomes cellulaires dans la particule "par accident". Ces ribosomes ne sont pas impliqués dans la synthèse des protéines virales. |
||||||
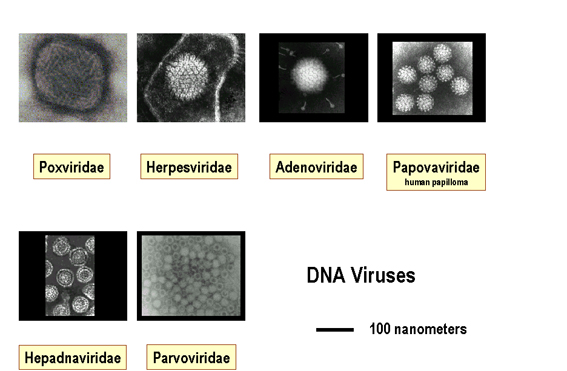 ADN
ADN
 ARN, +brin
ARN, +brin
 ARN, -brin
ARN, -brin
Images © 1995 Dr
Linda Stannard, University of Cape Town and © 1994 Veterinary Sciences Division,
Queen's University Belfast
Figure 2. Taille relative de virus à ADN et virus à ARN
Les virus infectent tous les principaux groupes d'organismes: les vertébrés, les invertébrés, les plantes, les champignons, et les bactéries, mais certains virus ont une gamme d'hôte plus large que d'autres. Par contre, aucun virus ne peut traverser la frontière eucaryotes / procaryotes.
Les facteurs qui influent sur la gamme d'hôte comprennent:
Est-ce que le virus est capable d'entrer dans la cellule hôte? C'est à dire, possède-t-il la protéine d'attachement qui le permet de se lier à un récepteur à la surface cellulaire? Par exemple, le VIH est largement restreint aux cellules qui expriment l'antigène CD4 à la surface (comme les lymphocytes T CD4+).
Dans le cas où le virus rentre dans la cellule, est-ce que la machinerie cellulaire adéquate est disponible pour la réplication virale? Par exemple, certains virus à ADN, comme le parvovirus B19, ne peuvent se reproduire que dans les cellules en division, qui ont des niveaux suffisamment élevés de désoxyribonucléotides pour la synthèse de l'ADN viral.
Dans le cas où le virus peut se répliquer, est-ce que des particules virales infectieuses peuvent quitter la cellule et propager l'infection? Par exemple, pour rendre la particule virale infectieuse, l'hémagglutinine du virus de la grippe saisonnière doit être clivée par une protéase cellulaire. Seules les cellules qui expriment ces protéases sont capables de propager l'infection virale.
Structure des
Particules Virales
Les virus varient en taille de 20 nanomètres de diamètre, comme les
Parvoviridae, à plusieurs centaines de nanomètres de longueur dans le cas
des Filoviridae (Figures 1 et 2).
Tous les virus contiennent un génome d'acide nucléique (ADN ou ARN) et une
ou plusieurs protéines formant une couche protectrice, appelée la capside.
L'ensemble formé par l'association entre la ou les protéines virales de
capside, et le génome viral est appelé la nucléocapside, qui peut avoir une
symétrie hélicoïdale, icosaédrique, ou complexe. Si la particule virale est
composée uniquement de protéine et d'acide nucléique, on parle alors d'un
virus nu ou non-enveloppé.
En revanche, les virus enveloppés sont dotées d'une enveloppe en bicouche
lipidique qu'ils obtiennent par bourgeonnement à travers une membrane de la
cellule hôte. Dans certains cas, les virus bourgeonnent à travers la
membrane plasmique, mais dans d'autres cas l'enveloppe peut être dérivée des
membranes cellulaires internes, telles que les membranes de l'appareil de
Golgi ou le noyau. Certains virus bourgeonnent à des sites spécialisés de la
membrane plasmique de la cellule hôte. Par exemple, le virus Ebola s'associe
avec des radeaux lipidiques qui sont riches en sphingomyéline, de
cholestérol et en protéines fixées par un ancre GPI (glycophosphatidylinositol).
Puisque le processus de bourgeonnement n'est pas obligatoirement létale pour
la cellule, la libération des particules virales enveloppés n'est pas
toujours accompagnée de la mort de la cellule hôte. Certains virus
enveloppés peuvent donc instaurer des infections persistantes.
Les virus enveloppés sont infectieux seulement si l'enveloppe est intacte,
car les protéines d'attachement virales qui reconnaissent les récepteurs de
cellules hôtes se trouvent dans l'enveloppe virale. Par conséquent, les
agents qui attaquent l'enveloppe, tels que les alcools et détergents,
suppriment l'infectivité des virus enveloppés. Pour cette raison les virus
enveloppés sont, de façon générale, plus facile à inactiver que les virus
non-enveloppés.
WEB RESOURCES
Principles
of virus architecture
Linda Stannard
Platonic Solids
University of Utah
Triangulation Numbers
J-Y Sgro
Structure des nucléocapsides virales
Symétrie icosaédrique
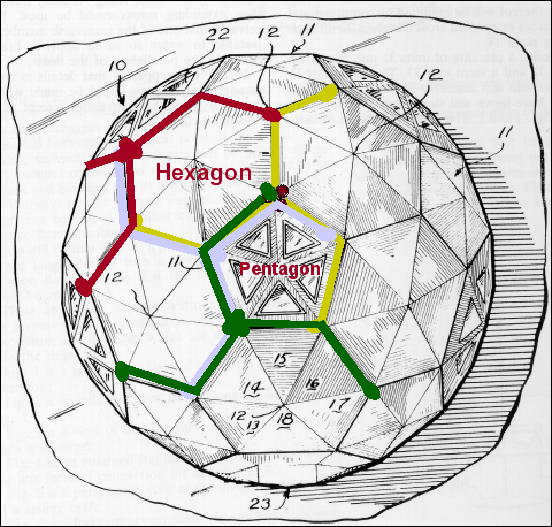
Un icosaèdre est un solide platonique comportant vingt faces triangulaires (figure 3A). Les sommets d'un icosaèdre sont formés par la rencontre des pointes de cinq des faces triangulaires. Ainsi, les six axes de symétrie passant par les deux sommets opposés d'un icosaèdre sont d'ordre cinq. Les dix axes passant par les deux faces opposés sont d'ordre trois (parce qu'ils passent au centre d'un triangle équilatéral), et les quinze axes passant par le milieu de chaque arête sont d'ordre deux. C'est pour cette raison que l'icosaèdre est caractérisé par la symétrie de rotation 5:3:2 (figure 3B, C).
L'extérieur d'une capside virale
icosaédrique est composée de sous-unités répétées de protéine virale. Selon le
virus, une seule ou plusieurs protéines virales forment ces sous-unités. L'acide
nucléique est empaqueté à l'intérieur de la capside ou il est protégé de
l'environnement externe par la capside (figure 3D).
Les protéines de capside s'associent en unités structurelles,
appelées capsomères, que l'on peut voir en microscopie électronique, et qui
restent associées quand on commence à dissocier une capside par des techniques
biochimiques. Les capsomères peuvent être composés d'une ou de plusieurs
protéines virales. Les capsomères aux 12 sommets présentent une symétrie d'ordre
cinq, et chacun interagit avec 5 capsomères voisins. Ils sont donc désignés
comme des pentons ou pentamères (figure 3E). Plus une particule virale est
grande, plus elle contient de capsomères; les capsomères supplémentaires sont
disposées selon un réseau hexagonal régulier sur les faces des icosaèdres. Ils
ont six voisins et sont appelés hexons ou hexamères (figure 3F). La taille d'un
icosaèdre dépend de la taille et du nombre des capsomères. Il y aura toujours 12
pentons (un à chaque sommet), mais le nombre de hexons augmente avec la taille
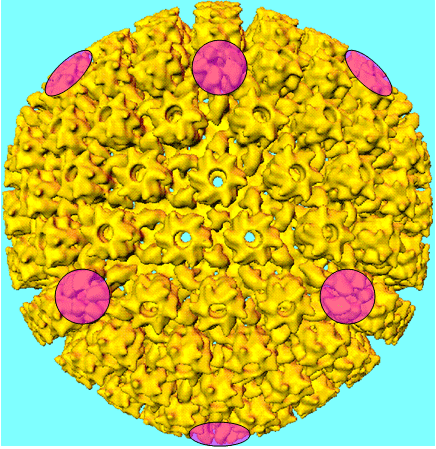
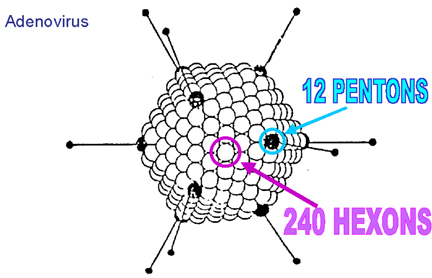
de la particule (figure 3H). Un bon exemple d'un virus icosaédrique est
l'adénovirus humain qui comprend les douze pentons obligatoires ainsi que 240
hexons (figure 3G et I).
La formation symétrique des matrices hexagonales sur une surface plane se
produit dans de nombreuses situations, par exemple, lors de l'emballage de tubes
à essai dans une boîte (figure 3J). Il peut également être décelé dans
l'organisation des sous-unités des protéines de capside du virus de l'herpès
simplex, un virus enveloppé icosaédrique. Dans la figure 3K, la membrane externe
du virus de l'herpès simplex a été éliminée afin de révéler la nucléocapside.
Bien que l'icosaèdre géométrique soit composé de faces plates (comme dans la
figure 3A), les particules virales icosaédriques sont habituellement sphériques
comme on le voit dans la figure 3K.
Un ballon de football (figure 3L), ou à une plus grande échelle, un dôme
géodésique (figure 3M) sont des exemples de structures sphériques composées de
pentons et de hexons, qui comme des particules virales, possèdent une symétrie
icosaédrique.
Figure 3
Capsides virales à symétrie icosaédrique
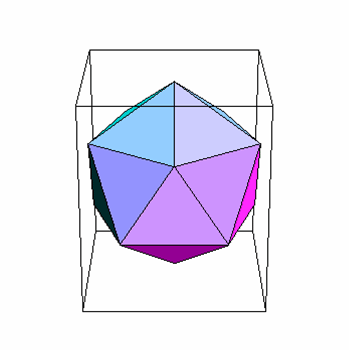 A: L'icosaèdre – une forme solide composée de 20 faces triangulaires
A: L'icosaèdre – une forme solide composée de 20 faces triangulaires
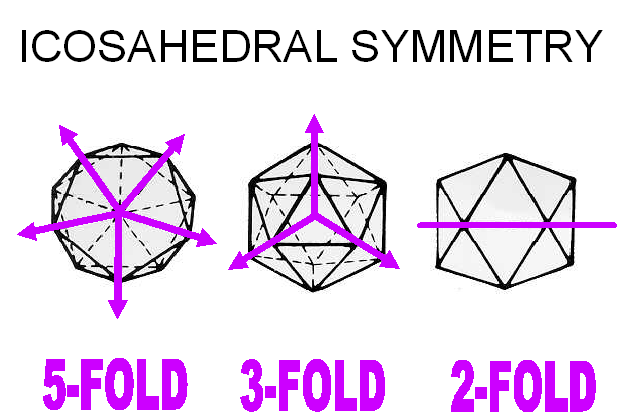 B: Symétrie axiale d'ordre 5:3:2
B: Symétrie axiale d'ordre 5:3:2
 C: Symétrie d'ordre 5 aux sommets
C: Symétrie d'ordre 5 aux sommets
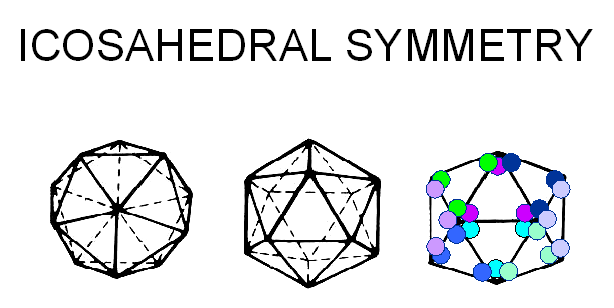
 D: L'acide nucléique est empaqueté à l'intérieur de la capside
D: L'acide nucléique est empaqueté à l'intérieur de la capside
© J-Y Sgro, Used with permission.
From
Virus World
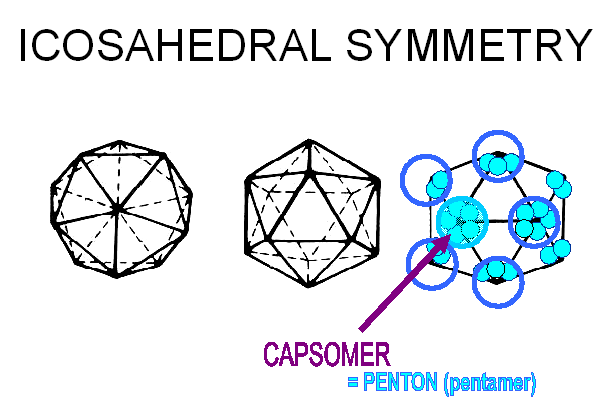 E: Les capsomères aux 12 sommets de l'icosaèdre ont une symétrie d'ordre 5, et
interagissent avec 5 capsomères voisins. Ils sont donc désignés comme des
pentons. (ou pentamères).
E: Les capsomères aux 12 sommets de l'icosaèdre ont une symétrie d'ordre 5, et
interagissent avec 5 capsomères voisins. Ils sont donc désignés comme des
pentons. (ou pentamères).
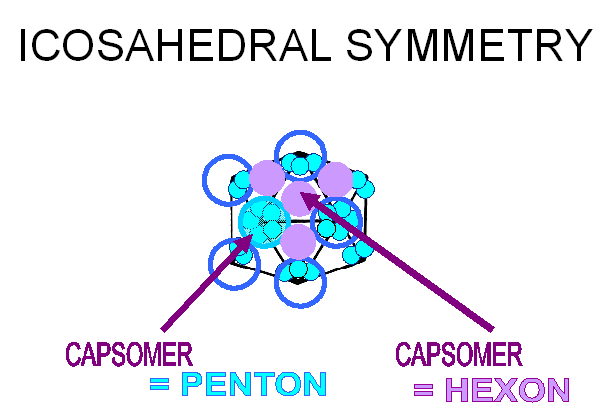 F-i: Plus qu'un virus est grand, plus sa capside contient de capsomères. Les
capsomères supplémentaires sont organisés en réseau hexagonal régulier sur les
faces de l'icosaèdre, et s'appellent des hexons.
F-i: Plus qu'un virus est grand, plus sa capside contient de capsomères. Les
capsomères supplémentaires sont organisés en réseau hexagonal régulier sur les
faces de l'icosaèdre, et s'appellent des hexons.
 F-ii: Nucléocapside d'un Herpesvirus. On peut voir des pentons au sommets de
l'icosaèdre.
Zhou et al. Baylor College of Medicine Reference: Z.
H. Zhou, B.V.V Prasad, J. Jakana, F.R. Rixon, W. Chiu Baylor College of
Medicine, Journal of Molecular Biology
F-ii: Nucléocapside d'un Herpesvirus. On peut voir des pentons au sommets de
l'icosaèdre.
Zhou et al. Baylor College of Medicine Reference: Z.
H. Zhou, B.V.V Prasad, J. Jakana, F.R. Rixon, W. Chiu Baylor College of
Medicine, Journal of Molecular Biology
 G: Symétrie d'une capside d'un Adénovirus
G: Symétrie d'une capside d'un Adénovirus
![]() H: Composants d'une capside icosaèdrique
H: Composants d'une capside icosaèdrique
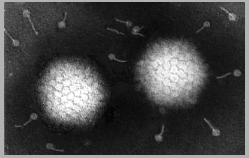 I: Adénovirus humain visualisé par microscopie électronique à coloration
négative
I: Adénovirus humain visualisé par microscopie électronique à coloration
négative
© 1995
Dr
Linda Stannard, University of Cape Town. Used with
permission
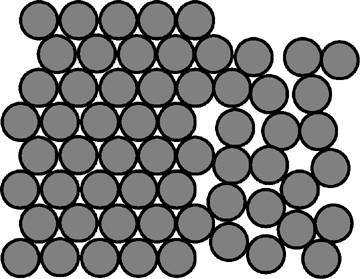 J: d'objets circulaires identiques dans un réseau hexagonal.
J: d'objets circulaires identiques dans un réseau hexagonal.
 K: Visualisation en 3 dimensions de la capside du
virus Herpes Simplex. La modélisation informatique a été effectuée à partir de
données de microscopie cryo-électronique.
National Institutes of Health
K: Visualisation en 3 dimensions de la capside du
virus Herpes Simplex. La modélisation informatique a été effectuée à partir de
données de microscopie cryo-électronique.
National Institutes of Health
 L: La structure icosaédrique d'un ballon de foot. A noter – l'associations de
pentons (en noir) et de hexons (en blanc) permet de former une structure
sphérique.
L: La structure icosaédrique d'un ballon de foot. A noter – l'associations de
pentons (en noir) et de hexons (en blanc) permet de former une structure
sphérique.
 M: Dôme géodésique.
M: Dôme géodésique.
Figure 4
Capsides virales à symétrie hélicoïdale.
Symétrie hélicoïdale
Dans le cas des capsides à symétrie hélicoïdale, les sous-unités protéiques de
la capside interagissent entre eux et avec l'acide nucléique pour former une
spirale. L'exemple classique d'un virus à symétrie hélicoïdale est le virus de
la mosaïque du tabac (figure 4 AE). La nature hélicoïdale de cette particule
virale est clairement visible en microscopie électronique à coloration négative,
car elle forme une structure rigide en bâtonnet. Chez les virus enveloppés à
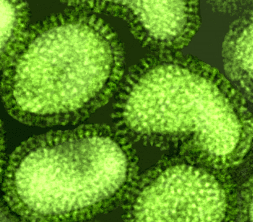
symétrie hélicoïdale (par exemple, le virus de la grippe, ou le virus de la
rage), la capside est plus souple et plus longue, et apparaît dans les clichés
de microscopie électronique plutôt sous l'aspect d'un cordon téléphonique
(figure 4H, I).
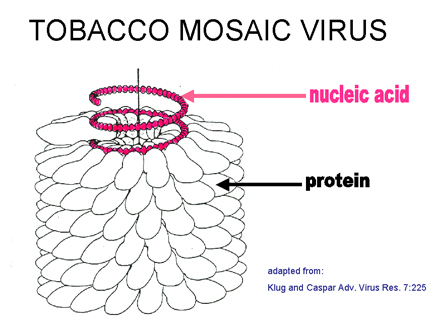 A: Symétrie hélicoïdale de la capside du virus de mosaïque du tabac (TMV)
A: Symétrie hélicoïdale de la capside du virus de mosaïque du tabac (TMV)
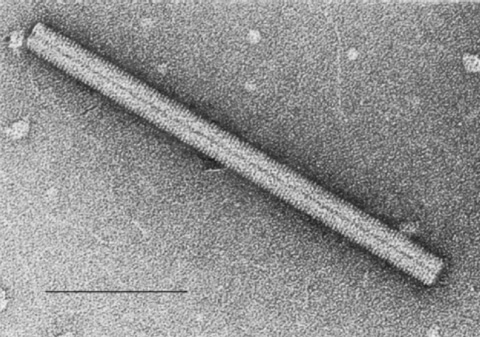 B: Image d'une seule particule virale du TMV en forme de bâtonnet par
microscopie électronique Image from the International Committee on Taxonomy of Viruses database.
B: Image d'une seule particule virale du TMV en forme de bâtonnet par
microscopie électronique Image from the International Committee on Taxonomy of Viruses database.
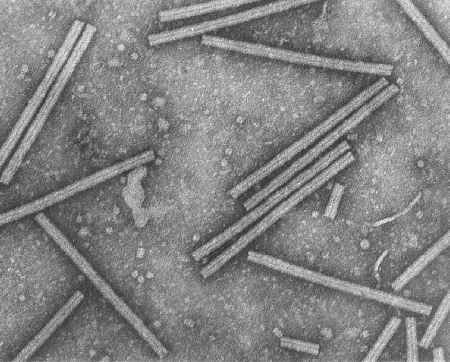 C Virus de mosaïque du tabac (TMV) vu par microscopie
électronique. © 1994 Rothamsted Experimental Station
C Virus de mosaïque du tabac (TMV) vu par microscopie
électronique. © 1994 Rothamsted Experimental Station
 D
Virus de mosaïque du tabac (TMV) vu par microscopie électronique. (TEM x207,480)
©
Dennis Kunkel Microscopy, Inc.
Used with permission
D
Virus de mosaïque du tabac (TMV) vu par microscopie électronique. (TEM x207,480)
©
Dennis Kunkel Microscopy, Inc.
Used with permission
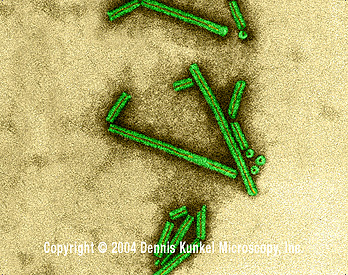 E
Virus de mosaïque du tabac (TMV) vu par microscopie électronique. (TEM x376,200) ©
Dennis Kunkel Microscopy, Inc.
Used with permission
E
Virus de mosaïque du tabac (TMV) vu par microscopie électronique. (TEM x376,200) ©
Dennis Kunkel Microscopy, Inc.
Used with permission
 F
Virus de la rage Wadsworth Center, NY Dept of
Health
F
Virus de la rage Wadsworth Center, NY Dept of
Health
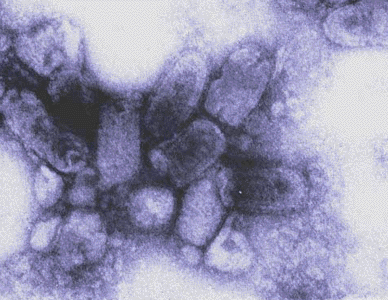
 G Virus de la grippe saisonnière vu
par microscopie électronique
© 1995
Dr
Linda Stannard, University of Cape Town. Used with
permission
G Virus de la grippe saisonnière vu
par microscopie électronique
© 1995
Dr
Linda Stannard, University of Cape Town. Used with
permission 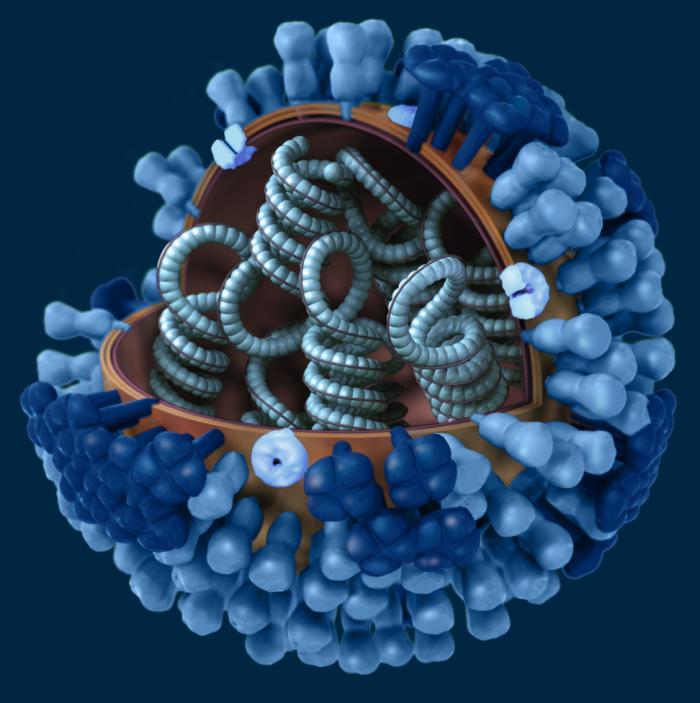 H Représentation graphique de l'ultrastructure d'une particule virale du
virus de la grippe. Une partie de l'enveloppe virale a été enlevée, ce qui
révèle la symétrie hélicoïdale des nucléocapsides à l'intérieur de la particule. CDC/ Doug Jordan
H Représentation graphique de l'ultrastructure d'une particule virale du
virus de la grippe. Une partie de l'enveloppe virale a été enlevée, ce qui
révèle la symétrie hélicoïdale des nucléocapsides à l'intérieur de la particule. CDC/ Doug Jordan
Symétrie Complexe
Certaines particules virales, comme celles
des Poxviridae, ne possèdent ni une symétrie hélicoïdale, ni une symétrie
icosaédrique. Leur structure est néanmoins régulière, et elles sont donc
désignées comme des particules de symétrie complexe. Récemment, il a été
démontré que les particules immatures des Poxviridae sont de forme sphérique,
avec une symétrie icosaédrique. Pour cette famille de virus au moins, la
structure complexe de la particule virale mature semble être dérivée d'une
structure icosaédrique.
Virus à structure complexe
 A Symétrie complexe des Poxviridae. Fenner and White Medical Virology 4th Ed. 1994
A Symétrie complexe des Poxviridae. Fenner and White Medical Virology 4th Ed. 1994
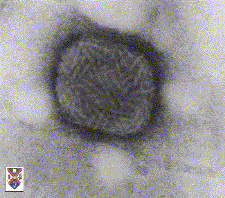 B
Des poxvirus visualisés par microscopie électronique.© Stewart McNulty, 1994
Veterinary Sciences Division,
Queen's University Belfast
Used with permission
B
Des poxvirus visualisés par microscopie électronique.© Stewart McNulty, 1994
Veterinary Sciences Division,
Queen's University Belfast
Used with permission
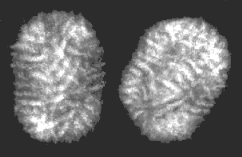 C
Molluscum contagiosum virus- a Molluscipoxvirus
© 1995
Dr
Linda Stannard, University of Cape Town. Used with
permission
C
Molluscum contagiosum virus- a Molluscipoxvirus
© 1995
Dr
Linda Stannard, University of Cape Town. Used with
permission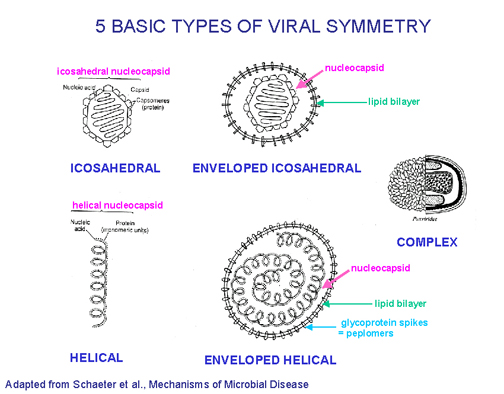 Les cinq formes de base des particules virales
Les cinq formes de base des particules viralesFigure 6
Les cinq formes de base des particules virales
- Icosaédrique, nu; par exemple, le virus de la poliomyélite, le virus de l'hépatite A (tous les deux des Picornaviridae), et les Adenoviridae
- Hélicoïdale, nu; par exemple, le virus du mosaïque du tabac. Ce type de particule virale est fréquemment retrouvé chez les virus des plantes, mais on le trouve chez aucun virus de l'Homme.
- Icosaédrique, enveloppé; par exemple, les Herpesviridae, le virus de la fièvre jaune (qui fait partie des Flaviviridae), et le virus de la rubéole (Rubivirus).
- Hélicoïdale, enveloppé; par exemple, le virus de la rage (Rhabdoviridae), le virus de la grippe (Orthomyxoviridae), le virus de la rougeole, et le virus des oreillons (tous les deux des Paramyxoviridae).
- Complexe; par exemple les Poxviridae
Les agents sub-viraux
Certains agents infectieux sont encore plus simple que des virus. En particulier, les satellites et les viroïdes possèdent des génomes en acide nucléique, qui ne codent pas pour une capside, tandis que les prions sont composés uniquement de protéine.
Satellites
Les satellites sont des molécules d'acide nucléique (ADN ou ARN) de 300 à 2000 nucléotides capables de se répliquer chez l'hôte uniquement en la présence d'un virus auxiliaire. Ils sont donc des parasites sur des virus.
Un satellite peut coder pour une protéine de capside, et dans ce cas, les cellules infectées par le satellite et son virus auxiliaire libèrent des particules composées de cette protéine et l'acide nucléique du satellite. Il est possible de distinguer les particules de ces virus satellites des particules du virus auxiliaire par microscopie électronique.
D'autres satellites ne codent pas pour une protéine de capside, mais empruntent les capsides de leur virus auxiliaire. Dans ce cas, les particules qui portent l'ARN ou l'ADN satellite sont identiques, ou très semblables aux particules du virus auxiliaire.
Viroïdes
Les viroïdes sont composés d'une molécule d'ARN simple brin, circulaire, de
moins de 400 nucléotides de longueur, avec une structure secondaire développée.
De façon étonnante, les ARNs des viroïdes ne sont pas encapsidés, et ne codent
pas pour des protéines, mais sont quand même capables de se répliquer et de
provoquer une pathologie chez leur hôte.
Le virus de l'hépatite delta
Les satellites et viroïdes se trouvent presque exclusivement chez les plantes. Cependant, au moins un agent semblable – le virus de l'hépatite delta (VHD) - est impliqué dans la pathologie humaine.
À certains égards, le VHD semble représenter une sorte d'intermédiaire entre un virus satellite et un viroïde. Son génome d'environ 1700 nucléotides comprend une partie riche en structure secondaire, qui ressemble à de l'ARN des viroïdes, mais une autre partie de l'ARN consiste en un cadre ouvert de lecture codant pour l'antigène de l'hépatite delta, qui sert de protéine de capside au VHD. Par contre, le VHD ne code pas pour sa propre protéine de fixation, qui doit être apporté par un virus auxiliaire. Dans l'occurrence, il s'agit du virus de l'hépatite B (VHB), un virus qui provoque des hépatites chroniques, et qui n'a aucun lien phylogénétique avec le VHD. Les particules infectieuses du VHD sont donc composées d'une nucléocapside associant l'ARN génomique du VHD et l'antigène de l'hépatite delta, entourée d'une enveloppe incorporant la glycoprotéine de fixation du VHB, l'antigène HBs.
Prions
Les prions sont composés uniquement de protéines – ce qui de premier abord semblerait incompatible avec leur réplication – car il n'existe aucun mécanisme biologique qui permet la lecture et la recopie d'une séquence d'acide aminés. La solution à ce paradoxe se trouve dans le fait que l'information génétique d'un prion ne consiste pas en sa structure primaire (sa séquence d'acides aminés) mais en sa structure trois dimensionnelle. En effet, des prions sont des formes “mal repliées” d'une protéine exprimée par l'hôte, capables d'imprimer leur conformation sur des copies "normales" de la même protéine. La protéine cellulaire ainsi transformée acquiert la capacité de transmettre l'information conformationnelle à d'autres copies de la protéine, et la répétition de ce processus propage l'infection. Des exemples de maladies à prion chez l'Homme sont le Kuru; la maladie de Creutzfeldt-Jacob et le syndrome Gerstmann-Straussler. Chez des animaux, on peut noter la tremblante du mouton, et la maladie de la vache folle.
Les virus sont-ils vivant ou morts?
La réponse à cette question dépend de la définition de la vie. D'un côté, les
virus font certainement partie du vivant, et partagent avec des organismes
vivant un génome en acide nucléique qui est transmis de génération en génération.
D'un autre côté, l'absence de métabolisme chez les virus, et en particulier leur
incapacité de fabriquer des protéines (due à l'absence de ribosomes chez les
virus) les distingue nettement de toute forme d'organisme cellulaire.
Curieusement, le fait que la question n'a pas de réponse évidente indique que la
frontière entre le vivant et le non-vivant n'est pas aussi nette que l'on
pouvait imaginer.
A cause de ce problème, il n'est pas d'usage d'utiliser le terme “virus vivant”,
quand on parle d'un virus infectieux, ni de parler de “tuer un virus”, quand on
cherche à l'inactiver par des moyens chimiques (exemple – par de la soude) ou
physiques (exemple – par la chaleur). Ainsi, dans un stock de virus, on peut
chercher à déterminer le nombre de particules virales infectieuses, et le nombre
de particules défectueuses (incapables d'infecter une cellule), et non pas le
nombre de particules virales “vivantes” et “mortes”.
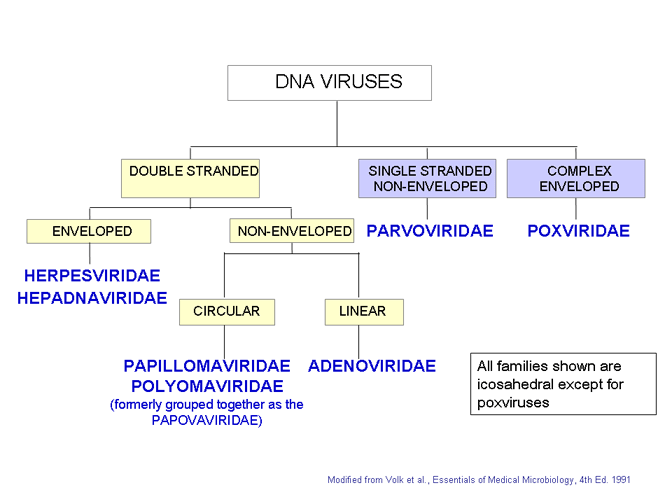 Les familles de virus à ADN. Toutes les familles sont montrées icosaèdre
sauf poxvirus
Les familles de virus à ADN. Toutes les familles sont montrées icosaèdre
sauf poxvirus
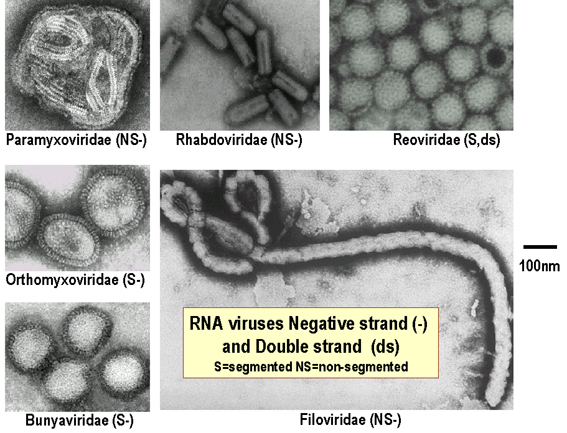
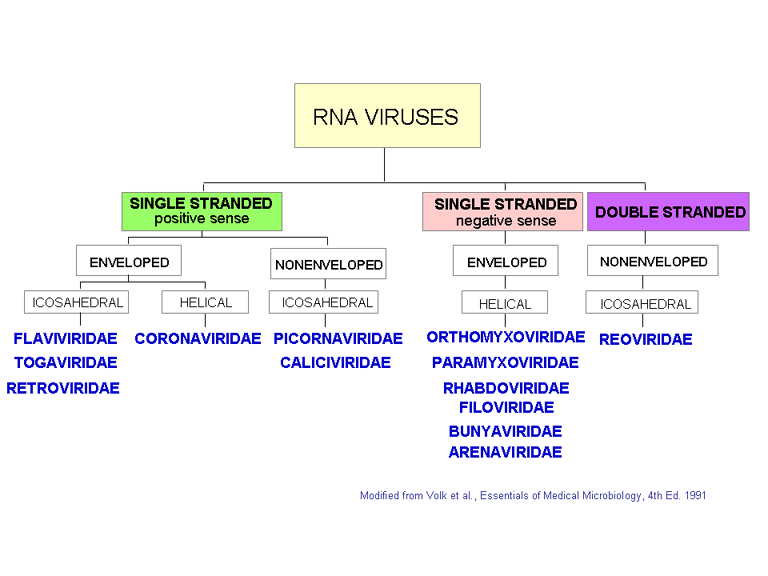 Les familles de virus à ARN
Les familles de virus à ARN
Figure 7
Modified from Volk et al., Essentials of Medical
Microbiology. 4th Ed
Classification des
Virus
Le système international de taxonomie virale est basé sur la structure et la
composition de la particule virale (Figure 7). Dans certains cas, le mode de
réplication est également utilisé comme critère de classification. Ainsi, les
virus sont classés en ordres (-virales), familles (-viridae), sous-familles (-virinae),
genres (-virus), et espèces. Par exemple, le virus de la rage fait partie du
genre Lyssavirus, de la famille des Rhabdoviridae et de l'ordre des
Mononegavirales.
Par contre, la plupart des familles de virus ne sont pas regroupées en ordres,
et il est également fréquent qu'une famille ne soit pas sous-divisée en
sous-familles. De plus, il existe un certain nombre d'espèces et de genres de
virus qui ne font pas partie d'une famille.
|
Caractères primaires utilisés dans la classification des virus |
|
|
Acide Nucléique |
ADN / ARN |
| Simple / double brin | |
| Segmenté / non-segmenté | |
| Linéaire / Circulaire | |
| Dans le cas d'un génome en ARN simple brin, s'agit-il d'un brin (+) qui peut fonctionner en tant qu'ARN messager, ou d'un brin (-) qui ne code pas directement pour des protéines virales. | |
| Génome diploïde (par exemple, chez les Retroviridae) ou haploïde dans la particule virale | |
|
Virion structure |
Capside Icosaédrique / Hélicoïdale / Complexe |
| Enveloppé / Nu | |
| Nombre de capsomères | |
Caractères secondaires
utilisés dans la classification des virus
Stratégie de réplication. Parfois, un groupe de virus défini par les critères
ci-dessus comporte un sous-groupe de virus qui emploient une stratégie de
réplication très différente de celle observée chez les autres virus du groupe.
Dans ce cas, le groupe sera divisé en fonction de la stratégie de réplication.
Dans un article intitulé "L'expression des génomes chez les virus d'animaux" publié en 1971, David Baltimore (Prix Nobel de Médecine 1975) a proposé un schéma de classification des virus au-dessus du niveau "Ordre", basé sur la composition de l'acide nucléique du génome viral, et sur les étapes majeures de la réplication et l'expression des génomes.
Ainsi, les rétrovirus, dont la particule renferme un génome en ARN, mais qui se répliquent via un intermédiaire en ADN au sein de la cellule infectée, forment une classe distincte dans la classification de Baltimore.
Les différentes classes de virus selon la classification de Baltimore sont:
| Classe de Baltimore | Génome viral | Caractéristiques | Polymérase dans la particule virale? |
| I | ADN double brin | Oui/Non | |
| II | ADN simple brin | Non | |
| III | ARN double brin | Oui | |
| IV | ARN brin (+) | Le génome viral code directement pour des protéines virales, comme un ARN messager | Non |
| V | ARN brin (-) | Le génome viral ne code pas directement pour des protéines virales | Oui |
| VI | ARN dans la particule virale, mais qui se répliquent via un intermédiaire en ADN au sein de la cellule infectée | La transcription inverse (molécule d'ARN recopiée en ADN) est nécessaire au cours de la réplication virale. | Oui |
| VII | ADN dans la particule virale, mais qui se répliquent via un intermédiaire en ARN au sein de la cellule infectée. | La transcription inverse (molécule d'ARN recopiée en ADN) est nécessaire au cours de la réplication virale. | Oui |
Les virus de classes VI sont les rétrovirus. Ils sont souvent regroupés avec les
virus de la classe VII, dans une catégorie qualifiée de virus à transcription
inverse.
Il est important de noter que les classes de Baltimore n'impliquent pas de lien
phylogénétique entre les virus au sein d'une même classe.
C'est à dire que, même si les virus d'une même famille (par exemple, les
Picornaviridae) semblent dérivés d'un ancêtre commun, il n'est pas exacte
d'affirmer que différentes familles de virus à génome ARN brin (+) (par exemple,
les Picornaviridae et les Caliciviridae) ont eu un ancêtre commun.
|
Classe I - Virus à Génome en ADN double brin |
|||||
|
|
Symétrie |
Enveloppe |
Taille (nm) |
Polymérase dans la particule virale? |
Commentaires et exemples |
| Papillomaviridae | I | - | 40-60 | - | Plusieurs papillomavirus humains (HPV) provoquent des verrues. Les génotypes HPV 16 et HPV 18 sont responsables du cancer du col de l'utérus. |
| Polyomaviridae | I | - | 40-60 | - | Le virus simien SV40 provoque des tumeurs chez les rongeurs. Son étude a permis de comprendre certains mécanismes moléculaires dans le développement des cancers. |
| Adenoviridae | I | - | 80 | - | Provoquent des rhumes et des gastroentérites. Développés comme vecteur viral pour la thérapie génique |
| Herpesviridae | I | + | 190 | - | Capables d'instaurer des infections latentes qui durent pendant plusieurs décennies. Exemples – virus de la varicelle; les virus Herpes simplex 1 et 2; le cytomégalovirus; le virus Epstein-Barr. |
| Poxviridae | C | + | 200-250 | + | Virus de la variole; de la vaccine. Réplication dans le cytoplasme de la cellule hôte. |
|
Classe II - Virus à Génome en ADN simple brin |
|||||
| Symétrie | Enveloppe | Taille (nm) | Polymérase dans la particule virale? | Commentaires et exemples | |
| Parvoviridae | I | - | 40 | - |
La plupart sont incapables de se répliquer de façon
indépendante – la coinfection par un virus auxiliaire (Adenovirus ou
Herpesvirus) est nécessaire. Développés comme vecteur viral pour la thérapie génique. |
|
Classe III - Virus à Génome en ARN double brin |
|||||
| Symétrie | Enveloppe | Taille (nm) | Polymérase dans la particule virale? | Commentaires et exemples | |
| Reoviridae | I | - | 75 | + |
La famille Reoviridae comprend les genres Orbivirus,
Reovirus, et Rotavirus. Les Rotavirus provoquent des gastroentérites. Les infections à Reovirus semblent asymptomatiques chez l'Homme, tandis que certains Orbivirus , comme le "Colorado Tick Fever Virus" sont pathogènes. Tous les Reoviridae ont des génomes segmentés. |
|
Classe IV - Virus à Génome en ARN brin (+) |
|||||
| Symétrie | Enveloppe | Taille (nm) | Polymérase dans la particule virale? | Commentaires et exemples | |
| Picornaviridae | I | - | 30 | - |
Plusieurs virus humains sont les Picornaviridae: Enterovirus, Rhinovirus (la plupart des rhumes communs), virus de la poliomyélite, le virus Coxsackie et le virus de l'hépatite A. Chez les animaux, le virus de la fièvre aphteuse fait partie des Picornaviridae. |
| Caliciviridae | I | - | 35 | - | Les Norovirus sont responsables des gastroentérites virales. |
| Flaviviridae | I | + | 40-55 | - | Comprend le virus de la fièvre jaune (flavus=jaune), le virus de la dengue et la virus de l'hépatite C. |
| Togaviridae | I | + | 60-70 | - |
Genre Alphavirus: le virus Chikungunya, le virus
Sindbis, le virus de la forêt Semliki, et différents virus provoquant
les encéphalites chez les équidés. Genre Rubrivirus: le virus de la rubéole. |
| Coronaviridae | I | + | 75-160 | - | Responsable de 10 à 30% des rhumes communs. Le syndrome respiratoire sévère aiguë (SRAS) est également dû à un Coronavirus. |
|
Classe V - Virus à Génome en ARN brin (-) |
|||||
| Symétrie | Enveloppe | Taille (nm) | Polymérase dans la particule virale? | Commentaires et exemples | |
| Rhabdoviridae | H | + | 60 x 180 | + | Virus de la rage, et le virus de la stomatite vésiculaire (pathogène chez le cochon, utilisé comme modèle pour l'étude de la réplication de Rhabdoviridae) |
| Paramyxoviridae | H | + | 150-300 | + | Plusieurs virus pathogènes: le virus de la rougeole, virus des oreillons, et le virus des syncytia respiratoire. |
| Filoviridae | H | + | 80 x 800-900 | + | Cette famille comprend des virus fortement pathogènes, comme le virus Ebola, ou le virus Marburg. Il s'agit de zoonoses – ces virus ne sont pas transmis de façon efficace entre humains. |
| Orthomyxoviridae | H | + | 80-120 | + | Génome segmenté – les virus de la grippe A et B |
| Bunyaviridae | H | + | 80-120 | + | Génome segmenté – plus de 80 membres, dont la plupart sont transmis par les arthropodes. Une exception sont les membre du genre Hantavirus, qui semblent être transmis par les rongeurs. |
| Arenaviridae | H | + | 60-130 | + | Génome segmenté – comprend le virus de la chorioméningite lymphocytaire (LCMV), un virus modèle important dans l'étude de la réponse immune anti-virale. |
|
Classe VI - Virus à rétrotransciption avec un génome en ARN |
|||||
| Symétrie | Enveloppe | Taille (nm) | Polymérase dans la particule virale? | Commentaires et exemples | |
| Retroviridae | I | + | 80-120 | + |
Deux virus pathogènes chez l'Homme: le HTLV-1 (virus avec
tropisme pour les lymphocytes T humain), et le VIH. Le génome est diploïde, c'est à dire que chaque particule virale renferme deux copies de l'ARN génomique. |
|
Classe VII - Virus à rétrotransciption avec un génome en ADN |
|||||
| Symétrie | Enveloppe | Taille (nm) | Polymérase dans la particule virale? | Commentaires et exemples | |
| Hepadnaviridae | I | + | 42 | + | Un seul virus pathogène chez l'Homme – le virus de l'hépatite B. L'infection peut mener à une hépatite B chronique progressive, qui est lié au développement du carcinome hépatocellulaire. |
|
|
|||||
![]()
Retour à la section de virologie de Microbiologie et Immunologie On-line
This page last changed on Wednesday, June 19, 2013
Page maintained by Richard Hunt