| x | x | |||||||||||||||||||||||||||||||
|
|
 |
|||||||||||||||||||||||||||||||
| BACTERIOLOGY | IMMUNOLOGY | MYCOLOGY | PARASITOLOGY | VIROLOGY | ||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||
| TURKISH | ||||||||||||||||||||||||||||||||
| SPANISH | ||||||||||||||||||||||||||||||||
| PORTUGUESE | ||||||||||||||||||||||||||||||||
| SHQIP - ALBANIAN | ||||||||||||||||||||||||||||||||
| FARSI | ||||||||||||||||||||||||||||||||
|
Let us know what you think |
||||||||||||||||||||||||||||||||
| SEARCH | ||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
| Logo image © Jeffrey Nelson, Rush University, Chicago, Illinois and The MicrobeLibrary | ||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
| KEY WORDS
Glycolysis |
Bacterial requirements for growth include sources of energy, "organic" carbon (e.g. sugars and fatty acids) and metal ions (e.g. iron). Optimal temperature, pH and the need (or lack of need for oxygen) are important. Oxygen Requirements Obligate aerobes must grow in the presence of oxygen; they cannot carry out fermentation. Obligate anaerobes do not carry out oxidative phosphorylation. Furthermore, they are killed by oxygen; they lack certain enzymes such as catalase [which breaks down hydrogen peroxide, H2O2, to water and oxygen], peroxidase [by which NADH + H2O2 are converted to NAD and O2] and superoxide dismutase [by which superoxide, O2., is converted to H2O2]. These enzymes detoxify peroxide and oxygen free radicals produced during metabolism in the presence of oxygen. Aerotolerant anaerobes are bacteria that respire anaerobically, but can survive in the presence of oxygen. Facultative anaerobes can perform both fermentation and aerobic respiration. In the presence of oxygen, anaerobic respiration is generally shut down and these organisms respire aerobically. Microaerophilic bacteria grow well in low concentrations of oxygen, but are killed by higher concentrations. Nutrient Requirements These include sources of organic carbon, nitrogen, phosphorus, sulfur and metal ions including iron. Bacteria secrete small molecules that bind iron (siderophores, e.g. enterobactin, mycobactin). Siderophores (with bound iron) are then internalized via receptors by the bacterial cell. The human host also has iron transport proteins (e.g. transferrin). Thus bacteria that ineffectively compete with the host for iron are poor pathogens. Temperature Bacteria may grow at a variety of temperatures from close to freezing to near to the boiling point of water. Those that grow best at the middle of this range are referred to as mesophiles; which includes all human pathogens and opportunists. (Those having lower and higher temperature optima are respectively known as psychrophiles and thermophiles). pH Many bacteria grow best at neutral pH; however certain bacteria can survive and even grow in quite acid or alkaline conditions.
|
|||||||||||||||||||||||||||||||
|
MOVIE Bacteria and Binary Fission |
||||||||||||||||||||||||||||||||
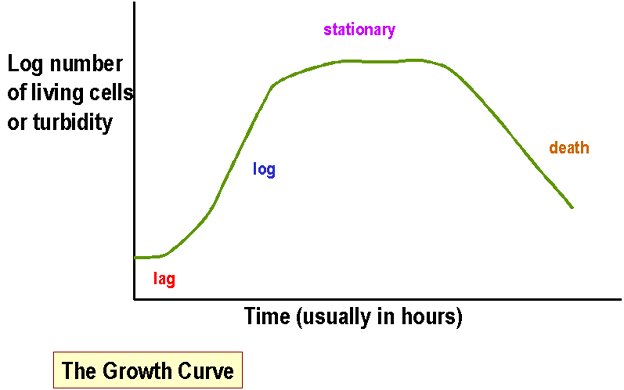 Figure 1. Growth Curve
Figure 1. Growth Curve |
Measuring bacterial mass in liquid cultures of bacteria
|
|||||||||||||||||||||||||||||||
|
|
METABOLISM OF SUGARS (as an example of metabolic pathways) Glycolysis (Embden, Meyerhof and Parnas [EMP] Pathway) This is the most common pathway in bacteria for sugar catabolism (It is also found in most animal and plant cells). A series of enzymatic processes result in conversion of sugars into pyruvate, generating ATP (adenosine triphosphate) and NADH (nicotinamide adenine dinucleotide). Chemical energy needed for biosynthetic purposes is stored in the newly formed compounds (ATP and NADH)
* refers to number of carbons in molecule
There are alternatives to this pathway for catabolizing sugars in order to produce stored energy within ATP. These include the pentose phosphate pathway (hexose monophosphate shunt) which is found in most animal and plant cells.. NADPH is generated using this pathway. Another pathway, the Entner Doudoroff pathway, is generally only found in certain bacterial cells Anaerobic Respiration Anaerobic respiration includes glycolysis and fermentation. During the latter stages of this process NADH (generated during glycolysis) is converted back to NAD by losing a hydrogen. The hydrogen is added to pyruvate and, depending on the bacterial species, a variety of metabolic end-products are produced.
Aerobic Respiration Aerobic Respiration involves glycolysis and the tricarboxylic acid cycle (Krebs cycle). Pyruvate is fully broken down to carbon dioxide (C1) and in the process, NAD is converted to NADH. Thus, in aerobic fermentation, NADH is generated from two sources (glycolysis and the Krebs cycle). Oxidative phosphorylation converts excess NADH back to NAD and in the process produces more ATP (stored energy). Ubiquinones and cytochromes are components of the electron transport chain involved in this latter process. Conversion of oxygen to water is the final step in the process.
|
|||||||||||||||||||||||||||||||
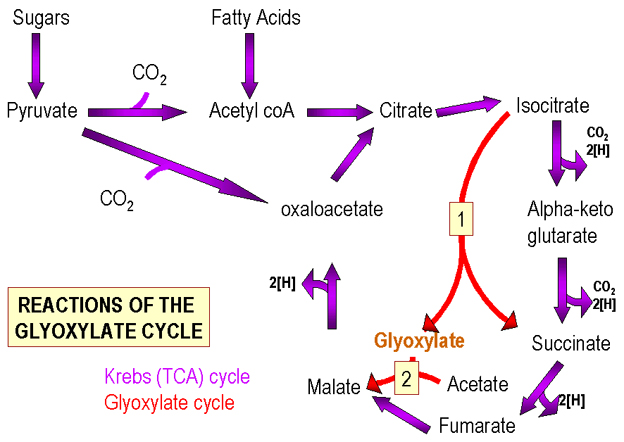 Figure 2. The Krebs and Glyoxylate Cycles
Figure 2. The Krebs and Glyoxylate Cycles |
Oxidative phosphorylation
The Krebs cycle (Figure 2) contains 4 and 6 carbon intermediates. Pyruvate (C3) can feed into the Krebs cycle in such a way that the number of C4/C6 intermediates remains the same or increases. a) Loss of CO2 (C1) from pyruvate to form acetyl coA, followed by addition to a C4 component of the cycle (oxaloacetate) produces a C6 component (citric acid). Thus the number of molecules of C6 produced equals the number of molecules of C4 initially present.
b) By addition of CO2 to pyruvate a C4 compound is produced. In this instance, additional molecules of C4 (a cycle component) are formed.
Thus if some of the cycle components are removed for use in other biosynthetic pathways, they can be replenished via this reaction.
|
|||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
|
METABOLISM OF FATTY ACIDS Fatty acids are broken down to acetyl groups (C2) which feed into the Krebs Cycle by addition to a C4 intermediate to produce C6. During the cycle, the added C2 is lost as CO2 and C4 regenerated. Overall, no increase in the number of molecules of cycle intermediates occurs. Thus, if fatty acids are the sole carbon source, no Krebs cycle intermediates can be removed without shutting it down:
Instead, bacteria utilize the glyoxylate cycle (Figure 2) (a modified Krebs Cycle) in which the enzymatic steps in which two CO2 molecules are removed from the C6 intermediate are by-passed. The C6 intermediate is converted to two C4 compounds (both components of the cycle). Thus for every acetyl group (from fatty acids), an additional cycle intermediate can be produced. The glyoxalate pathway is not generally found in animal cells since pre-formed fatty acids, present in the food, are utilized.
In summary, the Krebs Cycle functions in a biosynthetic and energy producing fashion. However, if intermediates are to be removed for use in other metabolic pathways, they must be replenished. The replenishment process for utilization of sugars and fatty acids is different.
|
|||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||